МОЛЕКУЛЯРНО-КЛЕТОЧНАЯ ПАЛЕОНТОЛОГИЯ НА 2007 ГОД: СВИДЕТЕЛЬСТВА О МАЛОМ
ВОЗРАСТЕ ЗЕМЛИ
Обзор
А.Н. Лунный
Доктор биологических наук, Москва
Lunarman@list.ru
THE MOLECULAR AND CELLULAR PALEONTOLOGY AT 2007: THE
EVIDENCES ABOUT SMALL AGE OF EARTH
Review
A.N. Lunarman
Ph.D.,
Moscow
РЕЗЮМЕ
В обзоре (264 ссылки) представлены наиболее полные в русскоязычной литературе сведения на начало 2007 г. из области молекулярно-клеточной палеонтологии (вопросы палеогенетики — древние ДНК — практически не затронуты). Рассмотрена история развития дисциплины, приведены данные о нахождении методами иммунохимии в костях динозавров и пр., возрастом в «миллионы — сотни миллионов лет» фрагментов белков, а также иммуногенных полипептидов.
«Достижением XXI века» являются работы М. Швейцер и др. (2005–2007; США) по обнаружению в костях ряда динозавров («65–80 млн. лет») гибких, прозрачных сосудов с морфологически цельными окрашенными эритроцитами, остеоцитов и костного матрикса. Рассмотрена идентификация в останках миоценовых амфибий («10 млн. лет») трехмерных структур костного мозга с клетками, сосудами, красным и желтым кровяными ростками и пр. (М. Макнамара и др., 2006; Ирландия). Представлены сведения о нахождении на кварцевых орудиях африканских гоминид («2 млн. лет») эритроцитов, лейкоцитов, жировых клеток и пр. (Т. Лой, 1998; Австралия).
Сравнительный анализ степени детекции фрагментов белков в палеонтологических («десятки — сотни миллионов лет») и археологических (тысячи — «десятки тысяч лет») образцах продемонстрировал отсутствие качественных отличий, несмотря на то, что оцененный возраст останков отличается на много порядков. Не выявлено закономерности при идентификации стабильного белка остеокальцина в костях, отличающихся по возрасту «в сотни раз».
Затронут радиационный аспект молекулярно-клеточной палеонтологии, связанный с оценкой доз облучения, накопленных ископаемыми биомолекулами и биоструктурами за приписанные им «миллионы — сотни миллионов лет» от внешних (радиационный фон Земли) и внутренних (поглощаемые ископаемыми костями уран и торий) источников. Сделано заключение, что столь большие получившиеся дозы радиации для морфологически неизменных сосудов, клеток и мягких тканей (в отличие от малых фрагментов белков) очень маловероятны.
Совокупность фактов в области молекулярно-клеточной палеонтологии на 2007 г. делает практически невозможным представление о длительных, многомиллионолетних геологических эпохах в истории Земли. Реальный период существования нашей планеты не оставляет времени для процесса макроэволюции, поэтому как дарвинистский, так и теологический эволюционизм (исповедуемый в том числе рядом православных России) несостоятельны.
СОДЕРЖАНИЕ
1. ВВЕДЕНИЕ
2. ПРЕДМЕТ «МОЛЕКУЛЯРНО-КЛЕТОЧНАЯ ПАЛЕОНТОЛОГИЯ»
3. СЛОЖИВШИЕСЯ ПРЕДСТАВЛЕНИЯ О ФОРМАХ СОХРАНЕНИЯ ИСКОПАЕМЫХ ОСТАТКОВ; ПСЕВДОМОРФОЗЫ И «МУМИИ»
4. СОХРАННОСТЬ БЕЛКОВ В ИСКОПАЕМЫХ ОСТАНКАХ
4.1. Теоретический размер определяемых с антителами фрагментов белков
4.2. Идентификация белков в ископаемых останках с оцененными возрастами в «миллионы — сотни миллионов лет»
4.3. Декларируемые механизмы сохранности белков в ископаемых останках
4.4. Сохранность белков в археологических, палеоантропологических и других остатках «малого» возраста
4.5. Остеокальцин: «чехарда» сроков сохранности
5. КЛЕТКИ И МЯГКИЕ ТКАНИ КАК ТАКОВЫЕ В ОСТАНКАХ ВОЗРАСТОМ В «ДЕСЯТКИ МИЛЛИОНОВ ЛЕТ»
5.1. Реальность опять невероятней фантастики
5.2. Первые относительно известные факты и их интерпретация
5.3. Публикации в «Science» 2005 г.: гибкие, прозрачные сосуды, эритроциты и остеоциты в костях четырех динозавров. А к 2007 г. — в костях еще нескольких
6. КОСТНЫЙ МОЗГ ВОЗРАСТОМ В «ДЕСЯТЬ МИЛЛИОНОВ ЛЕТ»
6.1. Самая нестойкая ткань может храниться с миоцена
6.2. Опрос патологоанатома
6.3. Гипотезы о сохранности костного мозга «миллионы лет»: кость как герметичный контейнер и самородная сера как стабилизатор
7. КЛЕТКИ, ОСТАТКИ КОСТНОГО МОЗГА, МЫШЦ, БЕЛКОВ И ПРОЧЕГО НА КАМЕННЫХ ОРУДИЯХ ВОЗРАСТОМ ДО «ДВУХ МИЛЛИОНОВ ЛЕТ»
8.
ОРГАНИЧЕСКИЙ ЗАПАХ ОТ ОКАМЕНЕЛЫХ КОСТЕЙ С АДСКОГО РУЧЬЯ
9. РАДИАЦИОННЫЙ АСПЕКТ МОЛЕКУЛЯРНО-КЛЕТОЧНОЙ ПАЛЕОНТОЛОГИИ
10. ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ.
Имея очи, не видите? имея уши, не слышите? и не помните? (Мк. 8:18)
1. ВВЕДЕНИЕ
Начиная с 2004 г. автор настоящего труда занимается проблемой идентификации биологических структур в ископаемых остатках[1]. Исследования являются только теоретическими, в форме научного анализа. Осуществляется сбор опубликованных фактов и их систематизация с последующими выводами, которые намного лучше укладываются в креационные, чем в эволюционные представления. В сборниках докладов на Рождественских чтениях, организуемых центром «Шестодневъ», издан ряд наших обзоров по молекулярной палеонтологии (2005–2007 гг.) [1–3]. В Интернете они первоначально были помещены на православном сайте «Слово отеческое» [4], а затем разошлись по Сети в качестве рефератов, предлагаемых студентам, и цитат-выдержек на различных форумах с сопутствующими обсуждениями. С конца 2006 г. все креационные труды автора данного обзора помещаются на страничку, выделенную ему в одной из популярных сетевых библиотек [5].
Представленный ниже труд включает в себя основные данные на конец 2006 г. — начало 2007 г. Ряд из них уже были разобраны нами в предыдущих статьях [1–3], но за время, прошедшее с момента их опубликования, появились новые факты. Некоторые сведения были обнаружены только сейчас. Кроме того, возникли новые аспекты интерпретации ранее приведенных фактов, а отдельные моменты, на которые уже обращалось внимание [3], нами, так сказать, «расширены и углублены». Некоторые же заключения уточнены и скорректированны.
Относительно возможности рецензирования данного труда, его автор должен сказать следующее. Судя по имеющимся публикациям, в России вряд ли есть специалисты, глубоко знакомые с данными по ископаемым белкам, клеткам и тканям в «миллионолетних» останках. Российская молекулярная палеонтология на настоящий момент исчерпывается, видимо, палеогенетикой — отдельными работами по выделению последовательностей древней ДНК возрастом максимум в «несколько десятков тысяч лет». Поэтому гипотетический рецензент нашего обзора должен будет дать себе труд сначала изучить не совсем знакомые ему проблемы.
Необходимо отметить также один момент, на который обычно обращают внимание научные работники — качество проанализированных в труде источников. Встречались замечания (на форумах в Интернете), что в предыдущих наших публикациях, дескать, многие источники — ненаучные. Но надо с полной ответственностью заявить, что основные рассмотренные нами факты по молекулярно-клеточной палеонтологии опубликованы именно в научной литературе, причем нередко — в ведущих мiровых журналах типа «Nature», «Science», «Proceedings of the National Academy of Sciences USA», «Biochemical and Biophysical Research Communication», «Geology» (США) и др. Конечно, молекулярная палеонтология — дисциплина достаточно узкая и необычная, поэтому ряд исследований увидели свет в гораздо менее известных журналах (с меньшими рейтингами и индексами цитирования), но и эти журналы — научные, многие — академические («Journal of Vertebrate Paleontology», «Journal of Protein Chemistry», «Palaios», «Applied Geochemistry», «PLoS Biology», «Naturwissenschaften», «Proceedings of the Royal Society B: Biological Sciences, London» и др.).
Из относительно необычного, что допускается в наших обзорах и что в принципе допустимо в настоящее время, можно назвать ссылки на официальные сетевые источники (сайты и электронные документы институтов и организаций). Примеры — Бюллетень университета в Северной Каролине, сайты Института динозавров в Монтане (Judith River Dinosaur Institute), университета в Квинсленде, сайт фирмы «Kodak» и пр. (дополнительные примеры можно найти в «Списке литературы»). Ныне одни научные журналы не допускают ссылки на Веб-источники, зато другие — допускают, правда — в зависимости от материала, на который ссылаются.
Однако в наших публикациях [1–3] (в том числе и в данной) есть ряд ссылок вспомогательного характера на научно-популярные издания («National Geographic News», «Discover» и т.п.) на источники в СМИ и даже на форумы. Здесь следует иметь в виду, что цель нашего креационного исследования лежит, помимо научной, еще и в идеологической сфере. Нам важно не только то, что опубликовано, но и то, как это интерпретируется в обыденно-научных представлениях[2], в научно-популярной литературе, в системе образования, в СМИ и в обществе.
Наконец, в обзоре есть, к примеру, научная ссылка на статью в «МК», объясняющаяся тем, что полноценная публикация об открытии в 2006 г. под Волгоградом окаменевшего мозга ископаемой птицы («90 млн. лет») пока отсутствует. Ссылки подобного рода здесь редки, а соответствующие данные носят весьма второстепенный характер. Они включены в материал только в познавательных целях (есть ли на свете окаменевший мозг мезозойской птицы, или же его на самом деле нет, на выводах нашей работы не отразится). Придираться к подобному подходу не следует — наша задача состоит не в написании канонической по форме научной работы, а в предоставлении, по возможности, всей последней информации по предмету. К тому же в «МК» применительно к окаменелому мозгу птицы помещено интервью с профессором Палеонтологического института РАН Е.Н. Курочкиным, имеющим значительный научный вес и множество соответствующих трудов (ссылки ниже).
2. ПРЕДМЕТ «МОЛЕКУЛЯРНО-КЛЕТОЧНАЯ ПАЛЕОНТОЛОГИЯ»
Молекулярно-клеточная палеонтология, ранее именуемая «молекулярной палеонтологией»[3] [6–8], — относительно молодая дисциплина. Ее официальная цель понимается различными исследователями по-разному. В соответствующих обзорах[4] имеются следующие определения:
· Изучение молекулярных следов жизнедеятельности организмов прошлого для выяснения по ним характера эволюции как живых организмов, так и биосферы в целом (член-корреспондент РАН А.Ю Розанов, директор Палеонтологического института РАН) [6].
· Выяснение событий прошлого путем исследования биомолекул и продуктов их долговременных превращений (диагенеза) в ископаемых источниках (ассистент профессора, доктор биологических наук Мэри Швейцер (M.H. Schweitzer) из университетов в Северной Каролине и в Монтане, ведущий молекулярный палеонтолог АН США, и член-корреспондент РАН В.А. Стоник из Дальневосточного государственного университета) [7, 8].
Достижение указанной цели (целей) предполагает решение следующих задач:
1. Изучение хемофосилий (биомаркеров) в осадочных породах для получения информации об изменениях климата, об источниках происхождении нефти и т.д. Известны, к примеру, следующие геобиомаркеры — продукты изменения метаболитов древних микроводорослей: диностеран, производное стеринового соединения диностерина, идентификация которого может пролить свет на те или иные особенности климата в прошлом, ботриококкан и каротан в некоторых месторождениях нефти, свидетельствующие о роли микроводорослей в ее образовании, и пр. [7].
2. Исследование в ископаемых останках вымерших животных и людей биологических макромолекул (белков, ДНК[5], липидов и пр.) [6–8]. Начиная с 2005 г. мы можем смело добавить к «биологическим макромолекулам» еще и «клетки и ткани» [2, 3]. Ибо, хотя кажется абсурдной сама мысль об обнаружении в «миллионолетних» костях структур, морфологически (по виду) и биохимически (органических по составу) идентичных клеткам и мягким тканям, ныне это стало реальностью (вопрос же о реальности самих «миллионов лет» мы пока не рассматриваем).
Если отойти от проблем, связанным с климатом и нефтью, то идеологической предпосылкой официально финансируемых работ в области молекулярно-клеточной палеонтологии и палеогенетики является попытка подкрепления макроэволюционных построений в первую очередь на неодарвинистском, молекулярно-генетическом, а теперь — даже на клеточном и тканевом уровнях. Проще говоря, заоблачная идеальная цель заключается в поиске эволюционных связок на молекулярно-клеточном и генетическом уровне между теми или иными классами и семействами животных. К примеру — поиск излюбленной гипотетической связки между динозаврами и птицами [9–11]. Или же, напротив, как в случае с людьми и «неандертальцами», — поиск отсутствия подобной связки [12, 13].
Целям же попроще, поприземленнее, посвящено, например, выяснение вопроса о родстве внутри того или иного рода или семейства. В результате пытаются определить, к какому слону ближе мамонт — к индийскому или к африканскому [14], и где находится место происхождения всего крупного рогатого скота Европы (оказалось — в разных местах [15]). Такие фундаментальные исследования, лежащие в рамках микроэволюционных представлений, не приносят, конечно, ничего эволюционно-сенсационного.
Начало молекулярной палеонтологии — момент достаточно темный, причем здесь не следует безоговорочно доверять самим палеонтологам, в том числе и «молекулярным». Истоком дисциплины считаются 1954–1956 гг., когда американец норвежского происхождения, физик, исследователь в области атомного оружия, доктор Филипп Абельсон (Philip Hauge Abelson) впервые опубликовал данные о выделении аминокислот из окаменелости девонской рыбы возрастом в «350 млн. лет»[6] [16]. Назвав эту дату в обзоре 2003 г. [8], доктор М. Швейцер далее переходит к 1974 г., когда была показана сохранность антигенных компонентов белков в «70 млн. лет» реакцией с антисывороткой [17].
Один из наиболее, как можно ожидать, известных палеонтологов России, директор палеонтологического музея РАН, член-корреспондент РАН А.Ю Розанов, в своем обзоре по современной палеонтологии за 1999 г. основоположниками молекулярного направления этой дисциплины почему-то называет Кальвина (не того, конечно) и В. Шопфа с соавторами, которые предложили какие-то «первые картины химической и биологической эволюции» [6]. Из публикации директора музея так и осталось не ясным, что же такого сделали упомянутые лица, кроме «картин».
В еще одном русскоязычном обзоре 2001 г., теперь уже конкретно по молекулярной палеонтологии, член-корреспондент РАН В.А. Стоник не называет никаких предтеч [7]. Мы можем добавить к этому, что в обеих цитированных отечественных публикациях в списках источников не приводится ни одна работа по собственно молекулярной палеонтологии белков из известных на тот период. Не упоминаются даже исследования ведущего молекулярного палеонтолога доктора М. Швейцер с соавторами, уже в 1990-х гг. накопивших ощутимый научный материал.
Вообще, все, что российские члены-корреспонденты РАН, знатоки предмета, относят к молекулярной палеонтологии животных, связано у них исключительно с исследованием ДНК в останках с оцененным возрастом максимум в несколько десятков тысяч лет. Для члена-корреспондента А.Ю. Розанова (как, впрочем, и для остальных указанных авторов) молекулярная палеонтология и палеогенетика (изучение древних ДНК) — синонимы, т.е. похоже, что этот автор в 1999 г. просто не имел представления о большинстве данных по молекулярной палеонтологии на тот период. В его обзоре [6] упоминается первое выделение ДНК из шкуры вымершего животного — квагги, истребленной в XIX в. (1984 г.; исток дисциплины «палеогенетики»). А также — из останков птиц моа, вымерших всего тысячу лет назад. Самый же древний образец в обзоре 1999 г. [6] представлен ДНК мамонта (как считается, мамонты вымерли от 10 до 50 тыс. лет назад).
Нам известен еще один соответствующий отечественный обзор, который был опубликован в самое последнее время — в 2006 г. в журнале «Генетика». Но он также полностью посвящен только древним ДНК [18].
Возможно, что с 2001 г. где-нибудь еще и увидели свет русскоязычные научные труды по молекулярно-клеточной палеонтологии ископаемых белков, клеток и тканей, помимо наших [1–3], но нам о них неизвестно. А в обзорах двух членов-корреспондентов за 1999 г. [6] и 2001 г. [7] никаких сведений подобного рода нет, хотя в позднем обзоре В.А. Стоника более ранний обзор А.Ю. Розанова и цитируется.
По нашему мнению, если не считать аминокислот 1954–1956 гг. [16][7], истинными пионерами молекулярно-клеточной палеонтологии являются польские авторы во главе с доктором Р Павлички (R. Pawlicki), с Факультета гистологии Ягеллонского Университета в Кракове. Еще в далеком 1966 г. ими были опубликованы данные об обнаружении в кости динозавра возрастом в «80 млн. лет» клеток, фибрилл коллагена и сосудов, причем опубликованы в одном из наиболее солидных мiровых журналов — «Nature» [19]. Следующая статья польских авторов увидела свет в 1972 г., здесь уже есть сведения о химическом анализе выявленных структур [20]. Всего с 1966 по 1998 г. доктором Р Павлички (в большинстве случаев без соавторов) было опубликовано не менее 15-ти научных трудов по изучению структур в костях динозавра, из которых 13 прямо относятся к области молекулярно-клеточной палеонтологии [19–31][8]. Большинство работ, правда, было опубликовано в относительно малоизвестных научных журналах.
Первоначально идентификация биоструктур проводилась польскими авторами под электронным микроскопом, и в костях динозавров были выявлены сосудистая система, фибриллы коллагена и остеоциты [19–21, 24, 25–27, 28, 29, 31], а в конце 1990-х гг. — даже эритроциты [31]. Кроме того, иммуногистохимическими методами идентифицировали ДНК, липиды и мукополисахариды (углеводы) в районах остеоцитов и, позже, путем рентгеноскопии — железо в эритроцитарных структурах [20, 22, 23, 26, 30, 31].
Необходимо отметить здесь, что примененные польскими авторами методы являются только косвенными. Они не использовали какие-либо специфические антитела, которые могли бы, например, однозначно указать на наличие коллагена или других белков хотя бы позвоночных в костях динозавра. Поначалу все ограничивалось изучением морфологии под микроскопом, когда о том, что открыты те или иные биоструктуры, судили по визуальным параметрам (грубо говоря, делали вывод: «очень похожи»). Либо, как в случае с эритроцитами, по тому факту, что, помимо «похожести», в районе их локализации имелся повышенный уровень железа. Использованные же Р.Павлички иммуногистохимические методы позволяли определить в срезах тканей (или костей) ДНК, липиды и углеводы, но отнюдь не доказать строго, что эти макромолекулы принадлежат именно динозавру, а не являются результатом посторонних загрязнений.
Поскольку польские авторы делали свои выводы по косвенным признакам и поскольку они опередили свое время применительно к молекулярно-клеточной палеонтологии, то их работам не уделялось достаточное внимание. В период, когда поздний И.А. Ефремов, профессор палеонтологии и мастер ископаемых находок, писал у нас свои последние фантастические романы, нигде даже мысли не допуская о нахождении в костях динозавров сохранившихся биоструктур, Р. Павлички с сотрудниками уже опубликовали соответствующую экспериментальную работу, да еще в «Nature» (1966) [19]. Получилось так, что реальность в очередной раз оказалась фантастичнее фантастики. Но только в XXI в., после аналогичных открытий более современными методами, выводы польских авторов стали действительно общепризнанными и бесспорными.
Признанными стали их выводы, но не статьи. Например, труды польских пионеров молекулярно-клеточной палеонтологии ведущим исследователем в этой области, доктором М. Швейцер, до 2005 г. не цитировались. Во всяком случае, в доступных нам статьях 1990-х гг. подобные факты обнаружены не были, равно как и в ее обзоре 2003 г. [8]. И только в 2005 г., в своей сенсационной работе по идентификации гибких прозрачных сосудов, окрашенных эритроцитов и остеоцитов в костях динозавров [32] (подробнее ниже), доктор М. Швейцер изволила наконец-то процитировать два труда Р. Павлички с сотрудниками [19, 31].
Первоначально мы обнаружили список некоторых публикаций польских авторов в статье креациониста Марка Армитейджа (Mark Armitage) [33], за что приносим ему благодарность. И мы из обзора в обзор продолжаем упоминать доктора Р Павлички как пионера указанной дисциплины [1–3], чтобы несколько исправить допущенную по отношению к нему научную некорректность.
* * *
В 1970-е гг., насколько известно нам и М.Швейцер [8], не считая пяти работ доктора Р. Павлички [20–24] было опубликовано порядка четырех исследований по молекулярной палеонтологии: помимо уже упоминавшихся антигенных структур (то есть остатков белков, реагирующих с антителами) возрастом в «70 млн. лет» (Нидерланды) [17], появились сообщения о выделении ощутимых по размеру фрагментов гликопротеинов (более 10.000 Да[9]) из раковины моллюска в «80 млн. лет» (США) [34] и идентификации антигенов также в раковине моллюска мелового периода (Нидерланды) [35]. Попытки же обнаружить полипептиды в окаменелостях колониальных морских организмов (граптолитов) возрастом в «450 млн. лет» не увенчались успехом — несмотря на значительную морфологическую и структурную сохранность, оригинальный биологический материал распался, заместившись, вероятно, минералами [36].
С начала 1980-х гг. наличие антигенных детерминант было показано уже для целого ряда образцов, включая коллаген и альбумин в костях возрастом до «1,9 млн. лет» [37, 38] и остеокальцин — в костях возрастом «13 и 30 млн. лет» [39]. Нечего и говорить о более «молодых» образцах: в 1980-е гг. остатки альбумина и коллагена были идентифицированы для «неандертальца», Homo Erectus, ископаемого бизона, мамонта [37, 38] и мастодонта (альбумин) [40]. Заметим здесь, что определяли не просто остатки белков как таковых, а остатки конкретных белков (альбумина, коллагена, остеокальцина), то есть, размер сохранившихся антигенных детерминант должен был быть достаточно ощутимым. Аналогичным образом обнаружили фрагменты полипептидов в останках исторических и доисторических людей (например, остатки гемоглобина в 1985 г. [41]). В 1980-х гг. появились первые обзоры по молекулярной палеонтологии [42, 43].
Короче, то, что даже сейчас кажется удивительным, стало почти обыденностью еще в 1980-х гг. А именно — факты обнаружения не полностью распавшихся за «биллиарды лет» фрагментов сложных и лабильных белковых макромолекул. Когда автор представленного вам труда узнавал все больше и больше подобных фактов из 1980-х — начала 1990-х гг. (причем оказывалось, что ни один из них за прошедшие десятилетия научной деятельности никогда не попадался на глаза), то постоянно вспоминался слоган из телевизионной рекламы: «А мужики-то — не знают!». Можно полагать, что если бы некоторые исследователи узнали бы про все такое ранее, то ряд вопросов был бы попросту снят уже в прошедшие десятилетия.
В 1990-х гг. молекулярно-клеточная палеонтология продолжала свое развитие, причем были достигнуты впечатляющие успехи, которым, однако, поверили далеко не сразу. Когда в 1997 г. доктор М. Швейцер с соавтором опубликовали данные об идентификации в кости тираннозавра эритроцитоподобных структур, то первоначально они увидели свет не в научном, а в научно-популярном журнале [44]. Но даже это вызвало тогда скандал (подробнее ниже; см. также в [1–3]). Про аналогичные же исследования польского доктора Р. Павлички еще 1966 г. [19] никто и не помнил, кроме, возможно, западных креационистов.
Мы не станем разбирать работы 1990-х гг., поскольку это было в основном сделано нами ранее [1], и, к тому же, соответствующие сводки данных представлены ниже в таблицах. Заметим здесь только, что качественный скачок в развитии молекулярно-клеточной палеонтологии был достигнут, все же, в последние годы, в XXI в.
* * *
Ископаемые ДНК. В настоящем обзоре мы не будем подробно рассматривать
данные палеогенетики, т.е. сведения об ископаемых ДНК. Начиная с 1984 г.,
когда впервые была получена ДНК вымершего животного (квагги) [6], в связи с
развитием метода полимеразной цепной реакции (PCR) предприняты сотни и даже тысячи попыток выделить
последовательности ДНК из древних и ископаемых источников. Многие такие попытки
оказались успешными. Количество публикаций в области палеогенетики несоизмеримо
с молекулярно-клеточной палеонтологией: уже в 1999 г. соответствующая
подборка литературы насчитывала более 1400 источников, а к 2004 г. была
составлена обширная энциклопедия статей по древним ДНК [45].
Начиная
с конца 1990-х гг. общепринятым в палеогенетике стало положение о том, что
ископаемая ДНК не может сохраняться
более нескольких десятков тысяч — максимум одной сотни тысяч лет в принципе,
исходя из уровней физико-химических воздействий в окружающей среде [1, 5, 6, 8,
46–48]. И если кто не знает прежних работ по палеогенетике, то он может
пребывать в радужном неведении об истинном положении дел (как, вероятно,
пребывают два члена-корреспондента РАН [6, 7]). В начале же 1990-х гг. с использованием метода PCR был проделан целый ряд работ, финансирование которых,
похоже, навеял «Парк Юрского периода» С. Спилберга. Из насекомых в янтаре,
из окаменелых растений и из ископаемых недр выделили и амплифицировали
последовательности ДНК и даже получили культуры
бактерий с оцененными возрастами от десятков
до сотен миллионов лет (в качестве обзора см. [48]). Наконец, тогда же почти
приблизились к идеалу С. Спилберга: извлекли фрагменты ДНК из останков
динозавров. В 1994 г. это проделали исследователи
из мормонского университета в Юте, США (кости возрастом «80 млн. лет»)
[49], а в 1996 г. —
пекинские авторы (яйцо ящера мелового периода) [50]. И если первая публикация известна
широко, причем удостоилась целого ряда критических комментариев [8, 48, 51], то
обсуждение второй, китайской, нам пока не встречалось.
Как
бы там ни было, но подобных работ с последовательностями ДНК возрастом в «десятки
и сотни миллионов лет» и с восставшими из чудовищных глубин времени бактериями
накопилось не менее чем десятка полтора — два, из разных стран и
лабораторий (ряд ссылок см. в [48]). Все они в настоящее время признаны
«неправильными», поскольку не соблюден ряд неких придуманных условий
достоверности. Среди последних гордо выделяется тест на так называемую
«эволюционную частоту». Он заключается в том, что, якобы, молекулярные «часы»
определяют время, за которое различные линии организмов, накапливая изменения в
своих ДНК, должны успеть «разойтись» друг от друга. Оценивается как бы
«эволюционное расстояние между последовательностями» и оно должно «соответствовать»
[48]. Мы пока что не станем углубляться в тонкости этого «научного
тестирования». Мы просто заметим, что если эволюционные расчеты времени, за
которое могли произойти наблюдаемые изменения в ДНК, не совпадают с оцененным
возрастом этих ДНК, то надо в первую очередь думать о том, что эволюционные
представления о возрастах не отвечают реальности. Те «миллионолетние» ДНК и
бактерии, что решили попросту выкинуть из накопленного научного багажа,
оттеняют субъективизм эволюционизма и представлений о длительных геологических
эпохах даже рельефнее, чем данные молекулярно-клеточной палеонтологии.
В
конце следует добавить, как и в нашем более раннем обзоре [1] мы отмечали, что,
вследствие крайней лабильности ДНК, ее чувствительности к посмертным изменениям
и к посторонним загрязнениям, вряд ли когда-нибудь удастся «клонировать
динозавра». Никакого «Парка Юрского периода» ожидать не следует, разве что по
типу империи Мавроди. Об этом говорят и ведущие специалисты по
молекулярно-клеточной палеонтологии (например, доктор М. Швейцер [8]).
3. СЛОЖИВШИЕСЯ ПРЕДСТАВЛЕНИЯ О ФОРМАХ СОХРАНЕНИЯ ИСКОПАЕМЫХ ОСТАТКОВ;
ПСЕВДОМОРФОЗЫ И «МУМИИ»
Исходя даже из физико-химических понятий, которые имеются у любого биохимика, молекулярного биолога или специалиста по биоорганической химии, кажется очевидным, что биологические макромолекулы, внутримолекулярные связи в которых характеризуются значительной свободной энергией, не могут быть устойчивыми в течение длительных периодов времени просто по определению, вследствие термодинамических процессов распада. Кроме того, внутримолекулярные связи должны окисляться в присутствии кислорода либо восстанавливаться в его отсутствии. Со ссылкой на палеонтологов из Университета в Северной Каролине (где работает доктор М. Швейцер), ранее было опубликовано утверждение, согласно которому по прежним представлениям органические молекулы в окаменелостях не способны сохраняться более 100.000 лет [46].
Важным кажется и воздействие микробиологического фактора: бактерии в течение многих тысячелетий — «миллионов лет» должны были разрушить все органические субстраты.
Российский автор обзора по молекулярной палеонтологии, член-корреспондент РАН В.А. Стоник отмечает [7], что макромолекулы претерпевают драматические посмертные изменения. Даже в тех редчайших случаях, когда тела живых организмов относительно хорошо сохранились морфологически, в них, согласно мнению В.А. Стоника 2001 г., можно найти только небольшие фрагменты биополимеров. Механизм распада связан со следующими факторами [7]:
· С автолизом, когда после гибели клеток их лизосомальные протеазы, нуклеазы и другие ферменты выходят вследствие разрушения мембран из мест своей локализации и расщепляют биомолекулы. Для автолиза нужна водная среда, поэтому в быстро подсыхающих участках, например в покровных тканях, он протекает с меньшей глубиной, чем в других органах и тканях.
· С перевариванием биомолекул микро- и макроорганизмами — трупоядами.
· С окислением/восстановлением макромолекул, если они захоронены в местах присутствия/отсутствия кислорода.
Член-корреспондент РАН В.А. Стоник указывает, что существенные изменения претерпевают не только сами исходные биополимеры, но и молекулы их вторичных метаболитов. Чаще всего они теряют функциональные группы и восстанавливаются. Если же захоронение подвергается воздействию высоких температур, то образуются конденсированные ароматические фрагменты [7]. Данный момент, как нам кажется, более касается месторождений нефти и газа, чем остатков ископаемых животных.
Несмотря на сказанное, палеонтологи сходятся в том, что скелетные системы некоторых вторичных метаболитов могут сохраняться в продуктах их долговременной трансформации в течение «многих миллионов лет» [7, 8, 32]. К примеру, доктор М. Швейцер с соавторами в [32] отмечают, что для окаменелых образцов возможна исключительная морфологическая сохранность, включая перьеподобные структуры [9, 52, 53], шерсть [54], цветность или цветные образцы [54, 55], эмбриональные мягкие ткани [56], мышечные ткани и/или внутренние органы [32, 57, 58], а также клеточные структуры [19, 31, 54, 59]. Правда, далее в [32] указано, что мягкие ткани сохраняются обычно не в виде тканей как таковых, а в виде углеродистых отпечатков (carbon films) [52, 53, 57] или в виде перминерализованных[10] трехсторонних реплик [56, 58, 60].
Ранее для сохранившихся ископаемых остатков было постулировано их окаменение с постепенным замещением органических структур неорганическими соединениями (с низкой свободной энергией связей, а потому — стабильными). По мнению, разделявшемуся палеонтологами в течение лет ста пятидесяти, ископаемые остатки древнего возраста должны быть представлены только в виде окаменелостей, отпечатков, хитина, панцирей и т.п., причем в окаменелости переходят не только скелеты, но и мягкие ткани.
Процессу окаменения (fossilize; фоссилизации) кроме остатков животных могут подвергаться также остатки растений, когда их ткани, в частности древесина, замещаются кремнеземом. При нехватке кислорода стволы и стебли растений превращаются в уголь, а листья — в углеродистые пленки. Аналогичным процессам фоссилизации иногда подвергаются микроорганизмы, но это происходит в исключительных условиях, когда их остатки захороняются и консервируются в тонкодисперсных осадках либо в коллоидных отложениях кремнезема [61].
В результате могут формироваться псевдоморфы, или псевдоморфозы, т.е. минеральные образования, обладающие внешней формой, необычной для слагающего их вещества, которые морфологически имитируют исходные органические структуры[11]. О псевдоморфозах за счет коллоидов земной коры писал еще В.И. Вернадский [62]. Бывают удивительные образования. Некогда в железорудной шахте погиб рудокоп. И шестьдесят лет спустя шахтеры обнаружили под землей человека, сплошь состоящего из сернистого железа. Данный псевдоморфоз вошел в историю под названием «пиритовый человек» [62]. Нередки также псевдоморфозы древних деревьев, причем в твердом минерале, заменившем с годами древесину, сохраняется даже микроскопическое клеточное строение [61, 62].
В середине 1980-х гг. музей Креационных свидетельств (The Creation Evidences Museum) приобрел интересный экспонат. В дорожном гравии из формации мелового периода был обнаружен псевдоморфоз — окаменелый человеческий палец (палец «Человека Мелового Периода»). После споров о том, действительно ли это палец или же нечто исходно минеральное, только похожее на него, была, наконец, проведена рентгеноскопия с компьютерной томографией, показавшей наличие внутри кости (рисунок 1) [63].
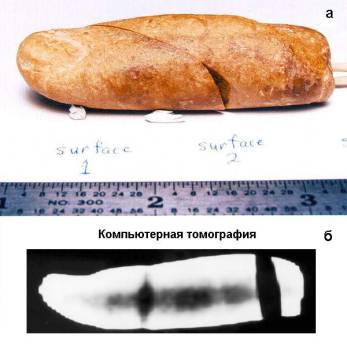
Рисунок 1. Найденный в гравии из меловых отложений псевдоморфоз
«Палец Человека Мелового Периода». а — общий вид; б — рентгеноскопия и компьютерная томография [63].
Редко встречающиеся палеонтологические «мумии» также являются не мумиями в прямом смысле (с мягкими тканями и с законсервированными органическими молекулами в их составе), а псевдоморфозами. Когда фоссилизованная кожа обернута вокруг скелета, когда образцы сохранили сколько-то внутренних тканей, то окаменелость рассматривается палеонтологами как мумия. Бывают мумии более или менее трехмерной формы, а бывают и относительно плоские (расплющенные в породе). Мумии же динозавров — это просто самые полные окаменелости подобных животных [64, 65]. Фоссилизованные чешуя, кожа и мягкая ткань найдены только у приблизительно одной из каждой тысячи окаменелостей динозавров, и обычно они охватывают менее одного процента образца [66]. Но встречаются мумии, у которых все это «охватывает» вплоть до 85–90% наружного покрова (см. ниже).
Всего на настоящий момент нам известно менее десятка мумий динозавров. В США в 2003 г. о них издана книжка с картинками, которая предназначена для детей, но весьма познавательна. В частности, там в порядке их обнаружения перечислены почти все мумии [67]. Первые три из известных мумий (утконосых динозавров, синоним — гадрозавры) были обнаружены Ч. Штенбергом (Ch. Sternberg) в 1908, 1910 и 1916 гг. в Вайоминге. Последний экспонат ушел на дно в северной Атлантике вместе с потопленным транспортом, перевозившим его в Британский музей. Остальные окаменелости до сих пор экспонируются в Нью-Йоркском Американском музее естественной истории. До 2002 г. эти мумии считались наиболее полными для динозавров; у них сохранилась трехмерная структура и до 40% фоссилизованного покрова [68] (рисунок 2).

Рисунок 2.
Мумия гадрозавра, найденная Ч. Штенбергом в начале XX в. [67, 68].
Вплоть до 1980-х гг. был перерыв в зарегистрированных находках, зато за последние два с небольшим десятилетия найдены, вероятно, почти все остальные наиболее известные образцы. В таблице 1 представлена сводка данных об экземплярах «миллионолетних» ископаемых остатков с сохранившимися в результате псевдоморфозов окаменелыми мягкими тканями.
Таблица 1. Ископаемые остатки с фрагментами окаменелых мягких тканей (палеонтологические «мумии» и псевдоморфозы) с оцененным возрастом в «десятки и сотни миллионов лет»
|
Вид
организма; место находки |
Образец
окаменевшей ткани |
Год
находки или публикации |
Ссылки |
|
Три утконосых динозавра
(«мумии Штенберга»); Вайоминг, США |
Трехмерная структура и до
40% покровных тканей |
1908, 1910 и 1916 гг. |
[67–69] |
|
Динозавр
сципионикс (Scipionyx); Неаполь, Италия |
Фрагменты кишечника, печени,
некоторых мышц и дыхательного горла |
1983 г. |
[57, 67, 70] |
|
Тесцелозавр (Thescelosaurus); Южная Дакота, США |
Сердце |
1993 г. |
[71] |
|
Крылатый (feathered) динозавр
синозауроптерикс (Sinosauropteryx), Китай |
Фрагменты мягких тканей;
отпечатки тканей в грудной клетке |
1994 г. |
[53, 67] |
|
Титанозавр (Titanosaurus); Аргентина |
Эмбрион |
1998 г. |
[56, 67, 72] |
|
Тираннозавр (Tyrannosaurus) (от «65 млн. лет»); Вайоминг, США |
Фрагменты мягких тканей |
2000-е гг. |
[66] |
|
Утконосый динозавр (Brachylophosaurus); «мумия Леонардо» («77 млн. лет»); Монтана, США |
Сохранность тканей на
85–90% (кожа и чешуя; мышцы и внутренние органы, когти, клюв, «гребень»,
плечо, глотка, зоб, язык, подушечки лап и содержимое желудка) |
2000–2002 гг. |
[73] (см. также [66–68,
74–81]) |
|
Копролиты (окаменевшие
экскременты) тираннозавра (от «65 млн. лет»); США |
Мышечные ткани |
2004 г. |
[82] |
|
Брюхоногий моллюск из
силура («425 млн. лет») |
Трехмерная детальная структура
мягких тканей; пищеварительная система, мышца ноги, радула, рот и нога |
2006 г. |
[83] |
|
Колониальные морские организмы
граптолиты («450 млн. лет») |
Морфологическая и структурная
сохранность |
1972 г. |
[36]. |
|
Беспозвоночные из позднего
неопротерозоя и раннего фанерозоя (не менее «650 млн. лет» [46]) |
Эмбрионы (бластомеры) |
2006 г. |
[84] |
|
Личинка мухи из Антарктиды
(«3–17 млн. лет») |
Морфологическая
сохранность: видны дыхальца и пр. |
2003 г. |
[85, 86] |
|
Птица мезозоя энанциорнис
(«90 млн. лет»); Волгоградская обл., Россия |
Мозг |
2006 г. |
[87]* |
|
«Человек Мелового Периода»
(псевдоморфоз обнаружен в гравии, добытом из меловой формации; от
«65 млн. лет») |
Палец с видимой при рентгеноскопии
костью |
Приобретен креационным
музеем в середине 1980-х гг. |
[63] |
*
В «МК» [87] опубликовано интервью с профессором Палеонтологического музея РАН
Е.Н. Курочкиным, под руководством которого и было проведено исследование.
Этот автор известен своими трудами по изучению древних птиц и динозавров (см.,
к примеру, [88, 89]).
Из всего окаменелого, что включено в табл. 1, на первый взгляд наиболее впечатляют сохранившиеся «сотни миллионов лет» псевдоморфозы беспозвоночных. Можно еще привести пример окаменелого червя, найденного в графстве Сомервел (Somervel), Техас [63] (рисунок 3).

Рисунок 3. Окаменелый червь из графства Сомервел в Техасе [63].
Формирование таких уникальных окаменелостей, по-видимому, объясняется тем, что процесс фоссилизации не обязательно связан с очень длительными периодами времени: при определенном комплексе условий возможен достаточно быстрый переход органических соединений в окаменевший образец. С данным фактом соглашаются и эволюционисты, поскольку имеется ряд неопровержимых свидетельств. К примеру, это окаменевшее водяное колесо, погребенное в монолите скалы (фото помимо прочих источников см. в [90]), окаменевшие мешки с мукой и др. [33]. Кроме того, в фоссилизованных ископаемых тканях можно найти биомолекулы, что указывает на очень быстрое захоронение и минерализацию [72, 91, 92].
Самая знаменитая «мумия» ископаемых животных найдена в 2000 г., раскопана в 2001 г. и исследована в 2002 г. Это мумия утконосого динозавра брахилозавра (Brachylophosaurus) «Леонардо», возраст которой оценивают в «77 млн. лет». Она названа так по сохранившемуся неподалеку граффити «Leonard Webb and Geneva Jordan, 1917» [75]. Научных публикаций нами не обнаружено, за исключением доклада группы изучавших Леонардо исследователей на ежегодной встрече палеонтологов позвоночных в Окле, США, в 2002 г. [73]. Представленные ниже данные взяты из источников, включенных в соответствующую графу табл. 1.
Руководителем исследований Леонардо является Нейт Мурфи (Nate Murphy), куратор палеонтологии позвоночных из музея на Мальте, Монтана, и основатель Института динозавров (Judith River Dinosaur Institute). Леонардо обеспечил «звездный час» Нейту Мурфи.
Мумия на 85–90% сохранила окаменевший покров (кожа и чешуя), мягкие ткани и внутренние органы, когти, клюв и «гребень» (складку вдоль спины). Сохранились трехмерные «слепки» правой плечевой мышцы, ткани глотки, зоб, язык и подушечки трехпалых ног (рисунок 4).
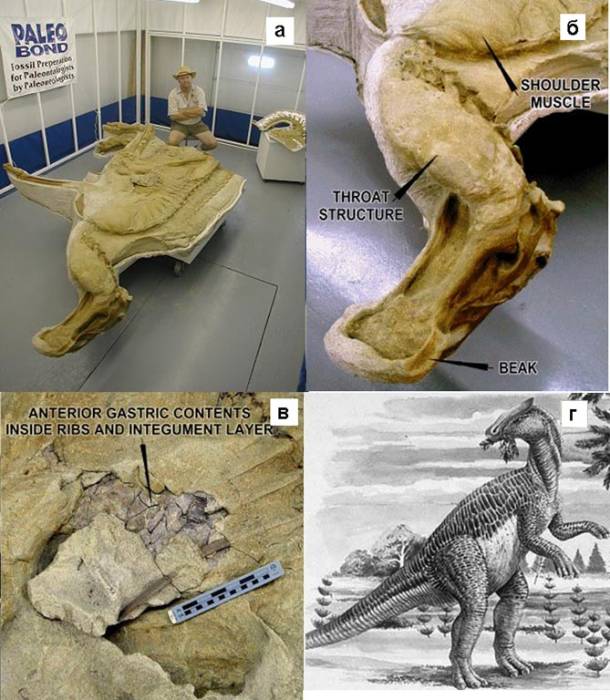
Рисунок 4. Мумия утконосого динозавра брахилозавра Леонардо. а —
мумия на стенде полевой станции Института динозавров; рядом Нейт Мурфи; б —
передняя часть с сохранившимися клювом, гортанью и плечевыми мышцами; в —
содержимое желудка внутри грудной клетки в месте случайного отпадения кожи и
наружных покровов [78]; г — реконструкция внешнего вида.
«Книгой рекордов Гиннеса» Леонардо был признан «Наиболее сохранившимся динозавром в мiре». На момент смерти он достигал семи метров в длину и весил около двух тонн. Экземпляр был полностью захоронен в песчанике, и чтобы не испортить экспонат, его извлекли целиком вместе с монолитом весом в 6,5 т (см. рис. 4, а).
Сохранилось даже окаменевшее содержимое желудка (рис. 4, в), так что стало возможным узнать состав последней трапезы животного: папоротники, печеночник[12], хвойные и магнолия. Желудок содержал пыльцу около 40 различных растений. Сохранившиеся ткани и облик вместе с содержимым желудка дают важную информацию о диете, способе передвижения (четвероногий или двуногий), локомоции динозавра и окружающей среде позднего мелового периода.
Мумию исследовали с помощью рентгеноскопии (компьютерная радиография), простого и оптического сканирования и компьютерной томографии, в результате чего были выполнены двумерные фотографии, по которым реконструировали трехмерное компьютерное изображение-модель. Внутри выявилось четырехкамерное сердце. Н. Мурфи даже представлял свои результаты на конференции по медицинским изображениям в Хьюстоне.
Образование больших фоссилизованных мумий, как и псевдоморфозов беспозвоночных (табл. 1; рис. 3), должно было быть процессом крайне быстрым, когда в течение нескольких дней нечто полностью останавливало разложение [81]. Поэтому в западных креационных кругах наличие подобных останков считают еще одним доказательством библейской катастрофы [80]. Так, скорее всего, оно и есть, но все-таки пока окаменелых мумий и псевдоморфозов обнаружено маловато, чтобы они могли служить твердым основанием для указанного заключения.
Полностью сохранившие внешний вид и форму внутренних органов окаменелости важны для реконструкции облика и строения ископаемых животных. Благодаря мумиям динозавров в последнее время попытались снять ряд вопросов, но эти исследования все еще не приобрели достаточную известность и никак не отразились на обыденно-научных представлениях. В частности, шея у утконосых динозавров оказалась не такой длинной, как реконструировали ранее и как рисовали и рисуют на картинках (ср. с рис. 4, а, б). Но самое, на наш взгляд, важное касается мумий сципионикса и синозауроптерикса (см. табл. 1). Останки детеныша динозавра сципионикса с фрагментами мягкой ткани были найдены возле Неаполя в 1983 г. [57, 67, 70]. Данная мумия предоставила свидетельства против гипотез о теплокровности динозавров и их родства с птицами: ободочная кишка локализовалась вблизи спинного хребта, как у крокодилов, но не как у птиц. Похожей на крокодилью оказалась и дыхательная система. В связи с этим палеонтологами-эволюционистами была выдвинута головоломная теория, что птицы возникли из крокодилов, которые когда-то эволюционировали от динозавров (ссылки см. в [70]).
«Мумия» крылатого китайского динозавра синозауроптерикса [53, 67] (см. табл. 1) также свидетельствует о различиях между динозаврами и птицами. Сохранившиеся отпечатки мягких тканей в грудной клетке по своей форме, а также по местоположению диафрагмы совпадают с тем, что наблюдается у крокодилов. У птиц же дыхательная система организована иначе — она не зависит от диафрагмы. Это означает, что птицы не могли произойти от животных, обладавших диафрагмой, так как две указанные дыхательные системы несовместимы [93].
Но в 2005 г. доктором М. Швейцер с соавторами при исследовании сохранившейся мягкой ткани в кости тираннозавра было, напротив, показано сходство между динозаврами и птицами. Оно заключалось в обнаружении у динозавра ткани медуллярной кости, которая из всех современных нам животных обнаружена только у самок птиц во время овуляции, но не у других яйцекладущих. Ее нет даже у крокодилов, которых часто относят к «потомкам динозавров» [11, 94].
Вот и смотрите, чему больше верить, результатам исследования «мумий» сципионикса и синозауроптерикса или костей тираннозавра. На наш же взгляд взаимопротиворечащие заключения связаны с тем, что динозавры — это динозавры, а птицы — птицы и есть, «вместе им не сойтись», о чем и свидетельствуют уверенно некоторые авторы [93–96].
Все, что мы рассмотрели в данном разделе, не касается напрямую сохранения биологических макромолекул и тканей как таковых в «миллионолетних» ископаемых остатках, хотя и нельзя уверенно отрицать их наличия в составе некоего каркаса в палеонтологических «мумиях» и псевдоморфозах, про которые полагают, что они полностью окаменели. Например, в материале из окаменевшей «мумии» эмбриона титанозавра в 2005 г. доктором М. Швейцер с соавторами были идентифицированы органические структуры и реагирующие с антителами фрагменты белков [72]. Выше уже отмечалось, что сведения о сохранности в окаменелостях остатков макромолекул получены достаточно давно [91, 92]. Так что те «камни», образовавшиеся в процессе диагенеза, которые представлены в табл. 1, может быть и не совсем камни по составу. Но такое их исследование еще впереди, в то время как для прочих ископаемых остатков за последние лет сорок уже получено множество действительно прямых данных.
4. СОХРАННОСТЬ БЕЛКОВ В ИСКОПАЕМЫХ ОСТАНКАХ
4.1. Теоретический размер определяемых с антителами фрагментов белков
Устойчивость белковой молекулы к
физико-химическим воздействиям зависит в первую очередь от ее структуры и
молекулярной массы. Низкомолекулярные белки, как правило, термостабильны, т.е.,
термодинамически устойчивы, что, конечно, продлевает им жизнь [97, 98]. Фибриллярные
белки, имеющие спиральную α-структуру
(α-кератин[13]
волос, шерсти, перьев, чешуи и т.п., коллаген[14])
более устойчивы, чем такие глобулярные белки, как альбумин, иммуноглобулин,
гемоглобин и пр. Особую же, порой чрезвычайную стабильность проявляют
полипептиды в жесткой β-конформации,
как, например, результат изменения исходного белка β-кератин [100] и вызывающие различные
заболевания («бешенство коров» и др.) патогенные формы прионов [101].
Понятно, что чем устойчивее в термодинамическом плане белок, тем он дольше сохраняется с течением времени. Это касается даже микробных воздействий, поскольку белки в жесткой β-конформации труднее перевариваются протеазами любого типа, в том числе и микробными [99–101]. И нам ясно, что, каковы бы ни были реальные, а не оцененные палеонтологами и геологами возрасты ископаемых остатков, пусть даже первые на три — четыре порядка меньше вторых, все равно белки шерсти, копыт, когтей и хрящей с соединительной тканью должны, в принципе, определяться лучше, чем альбумин, гемоглобин, гликопротеины и т.п. лабильные полипептиды.
Здесь надо уяснить себе, что значит «определяться». В большинстве ископаемых остатков, начиная с 1970-х гг., белки были детектированы радиоиммунологическими и/или иммунохимическими методами. Это связано с использованием, как правило, поликлональных антител в виде антисывороток. И возникает закономерный вопрос: каковы должны быть остаточные размеры фрагмента белка, чтобы он мог связываться с антителами? Точный ответ дать трудно (причем он зависит от структуры и свойств конкретного белка), но узнаваемые антителами фрагменты могут быть, в принципе, очень невелики по размерам. В то же время следует учитывать, что во многих случаях с помощью антисывороток были идентифицированы конкретные типы белков, а не просто какие-то остатки с пептидными связями. В ряде случаев, как мы увидим ниже, специфично определяли, к примеру, альбумин и коллаген по отдельности, отдельно детектировали древние иммуноглобулины гоминид и лошади и т.п. Это значит, что фрагменты белков в ископаемых остатках были настолько ощутимого размера, что могли сохранять элементы индивидуальной и даже видоспецифичной структуры конкретных полипептидных цепочек.
Трудно сказать однозначно, каков мог быть размер этих фрагментов. Примем, что масса средней аминокислоты составляет 140 у.е. (от 89 до 240; большинство 120–150) [100]. Пептиды, имеющие молекулярную массу менее 1000 (у.е.) (от 7–8 аминокислот), имеют слабую иммуногенность — способность вызывать образование антител [102, 103]. Участок же молекулы антигена (т.е., определяемого белка), отвечающий за связывание с антителом, содержит минимум 6–8 аминокислотных остатков, хотя, в реальности, этот участок (антигенная детерминанта; эпитоп) более определяется пространственной конформацией конкретного фрагмента белковой молекулы [104, 105]. Словом, в самом теоретически «худшем» случае размер идентифицированных в ископаемых остатках радиоиммунологическими и иммунохимическими методами пептидов не мог быть менее 6–8-ти аминокислот, но, учитывая указанную выше специфичность определения, должен был быть больше.
Что такое «цепочка из 6–8 аминокислот?» Молекулярная масса, к примеру, альбумина составляет 67.000 (при средней массе аминокислоты в 140 у.е. получается 479 остатков), а остеокальцина[15] — 5900 (точно 49 остатков) [97, 98]. Если принять указанный выше размер антигенной детерминанты (эпитопа) за 6–8 аминокислот, то получается, что для успешной идентификации альбумина в ископаемых останках в самом худшем случае должно было оставаться не менее 1–2% от интактного белка. Для остеокальцина же — не менее 12–16%.
Но не вся молекулярная палеонтология ископаемых белков исчерпывается только их качественной детекцией. Имело место и количественное выделение значительных по размеру фрагментов белков, и получение антител к экстракту из окаменелой кости.
4.2. Идентификация белков в ископаемых останках с оцененными возрастами
в «миллионы — сотни миллионов лет»
Наша подборка выполнена для образцов с принятым возрастом в «миллионы лет» и более. Нам интересны только такие сроки. «Мелочь» же, которая моложе (так, «тысяч до двухсот — пятисот лет»), будет частично рассмотрена ниже.
Кажется логичным разделить результаты исследования стабильных и лабильных белков. В таблице 2 представлены данные по идентификации в ископаемых образцах остатков кератина, коллагена и остеокальцина.
Таблица 2. Идентификация стабильных белков в ископаемых остатках с оцененными возрастами в «миллионы — сотни миллионов лет»
|
Белок |
Ископаемый
образец |
Оцененный
возраст |
Метод
определения |
Год публикации,
страна |
Ссылка |
|
β-Кератин |
Птица позднего мелового
периода Rahonavis ostromi |
Не менее 65 млн. лет |
Иммунохимия |
1999; США |
[106] |
|
Ящер мелового периода Shuvuuia deserti |
Не менее 65 млн. лет |
Иммунохимия |
1999; США |
[9] |
|
|
Коллаген |
Фрагменты черепа и плечевой
кости гоминид |
1,65 млн. лет |
Радиоиммунологический и иммуноферементный
(ELISA) анализы |
1997, 1998; Испания, США |
[107, 108] |
|
Останки австралопитека (Australopithecus robustus) |
1,9 млн. лет |
Радиоиммунологический
анализ |
1980, 1981; США |
[37, 38] |
|
|
Останки обезьяны Ramapithecus |
8 млн. лет |
Радиоиммунологический
анализ |
1983; США |
[109] |
|
|
Ископаемые кости |
10 млн. лет |
Иммуноблоттинг
(dot-blotting) |
1986 |
[110] |
|
|
Летучая мышь |
50 млн. лет |
Иммунохимия. Коллаген не определялся при морфологической
сохранности макроструктур |
2001; Австрия |
[111] |
|
|
Тираннозавр |
65 млн. лет |
Коллагеноподобные структуры
под электронным микроскопом |
2001 |
[33] |
|
|
Тираннозавр |
65–67 млн. лет |
Иммунохимия |
1994; США |
[112] |
|
|
Тираннозавр |
65–67 млн. лет |
Иммунохимия |
2005; США |
[32, 113] |
|
|
Птица позднего мелового
периода |
70 млн. лет |
Иммунохимия |
2005; США |
[114] |
|
|
Динозавр |
80 млн. лет |
Коллагеноподобные структуры
под электронным микроскопом |
1966; Польша |
[19] |
|
|
Ископаемая кость |
100 млн. лет |
Иммунохимия |
2002; США |
[115] (обзор) |
|
|
Утконосый динозавр Seismosaurus |
150 млн. лет |
Хроматография HPLC; одна из фракций соответствовала по параметрам
коллагену, но аминокислотный состав
отличался |
1991; США |
[116] |
|
|
Динозавр |
200 млн. лет |
Морфологически коллагеноподобные
структуры под электронным микроскопом |
1972 |
[117] |
|
|
Колониальные морские организмы
(граптолиты) |
450 млн. лет |
Иммунохимия. Коллаген не определялся при морфологической
сохранности макроструктур |
1972 |
[36] |
|
|
Остеокальцин |
Кости быка, зуб грызуна |
13 и 30 млн. лет
соответственно |
Очистка (HPLC и ионообменная хроматография) с существенным
выходом и аминокислотный анализ. По параметрам белки были неоличимы от
современного остеокальцина быка |
1987; Нидерланды |
[39] |
|
Тираннозавр |
65–67 млн. лет |
Иммунохимия |
2005; США |
[32, 113] |
|
|
Четыре динозавра юрского и
мелового периодов |
73–75 и 150 млн. лет |
Иммунохимия; хроматография HPLC; обнаружена специфичная для остеокальцина
аминокислота |
1992; Нидерланды |
[118–120] |
|
|
Игуанодон |
120 млн. лет |
Ионообменная хроматография
и иммунохимия |
2003; Великобритания |
[121] |
Из табл. 2 видно, что антигенные остатки стабильных белков свободно сохраняются в течение по крайней мере «десятков миллионов лет». Что касается коллагена, то его морфологические структуры, детектируемые под электронным микроскопом как коллагеновые тяжи, способны устоять и «сотни миллионов лет» [36, 115–117]. Но такие структуры могут являться результатом диагенеза, представлять собой некие псевдоморфозы, лишенные исходного органического материала. Так, в кости летучей мыши («50 млн. лет») [111] и в граптолитах («450 млн. лет») [36] иммунохимически коллаген не определялся. В останках же динозавра возрастом в «150 млн. лет», хотя коллагеноподобная фракция и имелась при хроматографии, однако ее аминокислотный состав сильно изменился в результате диагенеза.
Все это так, и, тем не менее, исходя из данных, представленных в табл. 2, мы можем сделать достаточно определенный вывод: антигенные детерминанты коллагена из разных источников свободно сохраняются в течение «70–100 млн. лет». Хотя, если отвлечься и встать на позиции физико-химической и биохимической логики, подобный сюрприз кажется маловероятным.
Но не станем отвлекаться — рассмотрим теперь остеокальцин, для которого, чтобы он был идентифицирован с помощью антител, должно сохраниться не менее 12–16% от исходной молекулы (немало, скажем прямо). Если взглянуть на табл. 2, то мы увидим, что в 1987 г. из останков возрастом в «десятки миллионов лет» был извлечен практически неизменный остеокальцин [39], что, опять же, невероятно с позиции обычной практики, несмотря на крайнюю термодинамическую устойчивость этого белка. А иммунохимически остеокальцин определялся уже для промежутков времени, больших, чем 100 млн. лет, в 1,2–1,5 раза [118–121]. Выходит так, что остеокальцин даже более устойчив к распаду во времени, чем коллаген, и это на настоящий момент самый «старинный» известный белок, хотя с его сроками сохранности в различных образцах и происходит какая-то «чехарда» (см. ниже).
Перейдем к идентификации нестойких, лабильных белков (таблица 3).
Таблица 3. Идентификация лабильных белков в ископаемых остатках с оцененными возрастами в «миллионы — сотни миллионов лет»
|
Белок |
Ископаемый
образец |
Оцененный
возраст |
Метод
определения |
Год публикации,
страна |
Ссылка |
|
Альбумин |
Фрагмент черепа и плечевой
кости гоминид |
1,65 млн. лет |
Радиоиммунологический и иммуноферементный
(ELISA) анализы |
1997, 1998; Испания, США |
[107, 108] |
|
Останки австралопитека (Australopithecus robustus) |
1,9 млн. лет |
Радиоиммунологический
анализ |
1980, 1981; США |
[37, 38] |
|
|
Останки обезьяны Ramapithecus |
8 млн. лет |
Радиоиммунологический
анализ |
1983; США |
[109] |
|
|
Иммуноглобулин G |
Кости гоминид |
1,6 млн. лет |
Иммуноблоттинг
(количественный dot-blotting); не было перекреста с антителами к
иммуноглобулину G лошади |
1998, 2002; Испания |
[108, 122] |
|
Кость лошади |
1,6 млн. лет |
Иммуноблоттинг
(количественный dot-blotting); не было перекреста с антителами к иммуноглобулину G человека |
2002; Испания |
[122] |
|
|
Трансферрин |
Фрагмент черепа гоминида |
1,6 млн. лет |
Радиоиммунологический и иммуноферементный
(ELISA) анализы |
1998; Испания |
[108] |
|
Гемоглобин |
Тираннозавр |
65 млн. лет |
Получение антисыворотки к
полипептидам экстракта из кости; реакция антисыворотки с современными
гемоглобинами (ELISA, иммуноблоттинг);
физико-химическое определение порфиринового железа |
1997; США |
[55] |
|
Гликопротеины |
Раковина моллюска |
80 млн. лет |
Разделение белков
гельфильтрацией; электрофорез в денатурирующих условиях и аминокислотный
анализ. Высокомолекулярные (более 10.000 Да) фракции соответствовали
остаткам гликопротеинов |
1976; США |
[34] |
|
Игуанодон |
120 млн. лет |
Разделение белков
ионообменной хроматографией; одна фракция соответствовала протеогликанам |
2003; Великобритания |
[121] |
|
|
Фосфопротеины |
Игуанодон |
120 млн. лет |
Разделение белков
ионообменной хроматографией; одна фракция соответствовала фосфопротеинам |
2003; Великобритания |
[121] |
|
Остатки белков |
Ископаемые останки |
70 млн. лет |
Реакция с антисывороткой |
1974; Нидерланды |
[17] |
|
Раковина моллюска мелового
периода |
Не менее 65 млн. лет |
Реакция с антисывороткой |
1979; Нидерланды |
[35] |
|
|
Эмбрион динозавра зауропода
позднего мелового периода |
Не менее 65 млн. лет |
Иммунохимия; антигенные
структуры были сходны с идентифицируемыми в современных источниках |
2005; США |
[72] |
|
|
Игуанодон |
120 млн. лет |
Выделение белков ионообменной
хроматографией; электрофорез; фракции с молекулярной массой от 5000 до
47.000–66.000 Да |
2000, 2003; Великобритания |
[121, 123] |
Логичны меньшие сроки, которые приписаны останкам с детектируемыми белками крови — альбумином [107, 108], иммуноглобулином G [108, 122] и трансферрином [108], хотя если сказать кому-нибудь из клинических биохимиков или судебных медиков, что подобные лабильные глобулярные белки не полностью распались за почти два миллиона лет, то вполне можно представить их реакцию. Но особенно прелестной является детекция альбумина в кости рамапитека с оцененным возрастом в «восемь миллионов лет» [109]. Это было сделано в 1983 г., причем уже достаточно опытным в то время американским молекулярным палеонтологом из университета в Сан-Франциско Джерольдом Ловенштейном (Jerold M. Lowenstein) [37, 38, 42, 109], и он функционирует в этой области до сих пор, опубликовавшись, к примеру, в 2005 г. в соавторстве с доктором М. Швейцер [72].
Правда, сама соответствующая статья Дж.М. Ловенштейна 1983 г. [109] нам, конечно, недоступна, учитывая в том числе редкость журнала (нет даже реферата). Но мы имеем ее обсуждения в современных публикациях, на которые и опираемся. Приведем эти обсуждения дословно.
Из: [108] (1998 г.): «...it
has been possible using radioimmunoassay (RIA) to obtain useful phylogenetic information
from extinct species such as mammoths and mastodons, the South African quagga
(a zebra-like equid), the Tasmanian marsupial wolf, Steller’s sea cow, 8-Ma-old fossil apes from Pakistan,
and human remains that include an Egyptian mummy and Homo erectus (Lowenstein,
1981 [38], 1983 [109]; Lowenstein & Scheuenstuhl, 1991 (Обзор [124])».
Из [122] (2002 г.): «Lowenstein,
using a radioimmunoassay (RIA), detected species-specific collagen and serum factors in fossils as old as
a 0·5-Myr-old Homo erectus, a 1·9-Myr-old Australopithecus robustus (Lowenstein,
1981 [38]) and an 8-Myr-old Ramapithecus (Lowenstein,
1983 [109])».
К этому добавим, что в 1980 и 1981 гг. доктор Дж.М. Ловенштейн опубликовал результаты параллельных исследований альбумина и коллагена [37, 38] (см. табл. 2 и 3).
Подобные возрасты для идентификации альбумина в останках ископаемых гоминид, человекообразных обезьян и разных «дюжих» (robustus) австралопитеков (см. табл. 3) уже недвусмысленно свидетельствуют о степени «корректности» общепринятых ныне датировок.
Помимо альбумина, очень «хороши» также лабильные иммуноглобулин G и трансферрин в «1,6 млн. лет». Но особенно «хорош» был тот игуанодон в «120 млн. лет», для которого не просто идентифицировали остатки белков иммунохимически, а провели их экстракцию и разделение. И чего только не обнаружили там английские исследователи в 2000 и 2003 гг., вплоть до белков массой в 47.000–66.000 Да (см. табл. 2). [121, 123]. Тот факт, что не детектировался орнитин, продемонстрировало, как указано в [123], отсутствие бактериального загрязнения (т.е. считается, что белки были игуанодоновы).
Равным образом выделили впечатляющие по размерам полипептиды и из ископаемого моллюска в далеком 1976 г. [34]. И все подобное прекрасно подтверждается на эмбрионе зауропода в работе 2005 г. доктора М. Швейцер с соавторами, среди которых был и старый Дж.М. Ловенштейн [72]. По разным показателям свойства антигенных структур ископаемых полипептидов (не менее «65 млн. лет») не отличались от свойств современных нам белков [72].
И если для моллюска 1976 г. [34] еще как-то можно было предполагать, что он нечто вроде латимерии (хотя палеонтологи, наверное, тут обсмеют), то для игуанодона [121, 123] и яйца мелового динозавра [72] подобная ситуация невозможна априори. Либо нам придется считать, что палеонтологи путают с костями и яйцами игуанодонов и пр. останки и яйца каких-нибудь верблюдов, китов, бегемотов или страусов. Мы такую возможность отбрасываем с негодованием.
В результате получается странная вещь: если для фрагментов стабильных белков в ископаемых останках показана максимальная сохранность в течение «десятков — сотни миллионов лет» (табл. 2), то и для лабильных, растворимых белков — тоже от «8 до 120 миллионов лет». Причем для растворимых ситуация даже круче — полипептиды в «70 и 120 млн. лет» выделены и охарактеризованы биохимически, а их размер составил вплоть до 10.000–66.000 Да [34, 121], что отнюдь не «1–2%» сохранившихся антигенных детерминант (см. выше подраздел 4.1), а гораздо больше, если не все 100%.
Ранее, в своем первом обзоре по молекулярной (тогда) палеонтологии [1], мы особое внимание уделили опытам доктора М. Швейцер с соавторами по идентификации в «недоокаменевшей» кости тираннозавра остатков гемоглобина [55]. Эти эксперименты несколько выделяются среди прочих, представленных в табл. 2 и 3, по своему методическому подходу. Оказалось, что из кости тираннозавра в «65 млн. лет» можно выделить экстракт с полипептидами, которые сохранили иммуногенную активность, и при введении крысам индуцируют у тех образование антител. Данные антитела (антисыворотка) «узнавали» в ELISA и при иммуноблоттинге современные гемоглобины индюка, голубя и кролика. Однако при электрофорезе экстрактов из кости тираннозавра материал на электрофореграмме виден не был, т.е., полипептиды, все-таки, оказались небольшими. Учитывая это, а также минимальный размер остатков полипептидов, которые способны индуцировать образование антител (см. выше подраздел 4.1), мы тогда, в [1], пришли к выводу, что у тираннозавра от его исходной молекулы гемоглобина осталось не более 10–15% и не менее 3–5%.
Словом, какие бы подходы в области иммунохимических методов исследования ни использовались, во всех случаях следует, что от белков осталось порядка единиц — максимум десятка полтора процентов. Это если не считать тех моллюска в «70 млн. лет» и игуанодона в «120 млн. лет» (табл. 3). Они своими большими по размеру полипептидными остатками несколько смазывают закономерную картину того, какие фрагменты способны сохраняться.
В своем первом обзоре, который готовился в 2004 г. [1], я писал насчет того, что фрагменты гемоглобина тираннозавра не были видны при электрофорезе следующее:
«И скажем эволюционистам: для вас лучше не подкреплять, а то вдруг такие большие фрагменты белка выделят, что их даже при электрофорезе... видно будет... Что тогда о возрасте фрагментов думать станете?»
Тогда я видел, что из раковины моллюска еще в 1976 г. были все-таки выделены относительно большие по размеру белки (более 10.000 Да), которые можно детектировать и на электрофореграмме [34]. Но в то время я посчитал, что все эти моллюски мало отличаются от современных, и что с их датировкой просто в очередной раз произошла ошибка [1]. Наверное, они все-таки отличаются и все-таки вымерли. Статьи же про белки игуанодона [121, 123] в 2004 г., мною, к сожалению, были пропущены, и в обзоре [1] не рассматривались. Получилось так, что вся моя ирония и сарказм 2004 г., похоже, остались втуне: большие по размерам глико- и фосфопротеины, а также неколлагеновые белки, которые имеют массу до 66.000 Да и, следовательно, видны при электрофорезе как нельзя лучше, прекрасно выдерживают даже «120 млн. лет» [121, 123].
Что тут сказать... Только повторить, что реальность получается фантастичнее любой фантастики, если стоять на позиции «миллионов лет» возраста ископаемых остатков и длительности геологических эпох.
4.3. Декларируемые механизмы сохранности белков в ископаемых останках
Мы из обзора в обзор [1–3] уделяем данному вопросу специальный раздел. Не является исключением и настоящая публикация, хотя в целом ничего особенно нового не прибавилось. В основном же можно согласиться с мнением доктора М. Швейцер, которая после перечисления всех гипотез указывала в [8], что «механизм сохранения органических соединений (ДНК и белков) в течение геологических периодов времени еще предстоит выяснить».
Гипотетические механизмы сохранности биомолекул следующие [8].
1. Формирование в процессе распада и разрушения молекулярных связей во время диагенеза специфических биополимеров, устойчивых к дальнейшей деградации [125].
2. Стабилизация биомолекул через образование комплексов с органическими продуктами перегноя в почве, в частности, с гуминовыми (гумусовыми) или с другими кислотами [7, 126]. Подобные комплексы, полученные и в лабораторных условиях, ингибируют активность расщепляющих ферментов [126, 127].
3. Быстрое «цементирование» при погребении отложений, что защищает от микробов. Этот процесс создает также закрытую систему, предотвращающую распад. Некоторые микроорганизмы могут формировать внеклеточные полимерные соединения на поверхности образцов, что способствует созданию изолирующей минеральной «пломбы» и окаменению. Подобные процессы приводят к сохранению макро- и микроструктур, иногда даже мягких тканей [128]. Данное явление связывают с формированием неких «внутренних кристаллов», предохраняющих белки и ДНК от внешних воздействий [8].
4. Связывание биологических макромолекул с минеральными субстратами, что повышает их стабильность. Этот механизм сохранения может быть наиболее важным. Максимального эффекта следует ожидать при его сочетании с формированием внутреннего закрытого «кристалла». Принято считать, что белки способны сохраняться очень долгое время, составляющее геологические эпохи, находясь в комплексе с апатитом минерализованных костей. Адсорбция на неорганической матрице предохраняет биомолекулы и от воздействия воды [8, 40, 43, 129–134].
Перечисленные механизмы, конечно, кажутся на первый взгляд вполне вероятными, в особенности последний. Вот только остается вопрос о тех реальных промежутках времени, в течение которых они способны действительно обеспечивать сохранение остатков белков, и вопрос о том, почему в реальности с возрастами сохранения одних и тех же белков наблюдается какая-то «чехарда» (см.ниже). А насчет «внутренних кристаллов» придется повторить [1, 3], что окаменелости — это все-таки не алмаз и даже не янтарь, чтобы миллионы лет сохранять внутри остатки белков, не пропуская ни микробов, ни кислоты почвы, ни кислород, ни радон, ни γ-излучение.
5. Специальный механизм повышения устойчивости гемоглобина через посредство свободных радикалов. Эти новые построения, развивающие упомянутую выше первую гипотезу, придуманы доктором М. Швейцер в самые последние годы [135]. Странная теория была доложена в 2006 г. на ежегодном форуме Американской ассоциации по продвижению науки (AAAS), одной из наиболее важных научных конференций в США, в виде презентации М. Швейцер и Дж. Хорнера (Jack Horner) [136]. Механизм, по мнению указанных исследователей, обусловлен железом в гемоглобине и миоглобине. После того как организм умирает, гемоглобин распадается, ионы железа освобождаются, становятся нестабильными, и при стабилизации генерируют окислительные свободные радикалы. Эти радикалы инициируют, де, формирование длинных молекулярных цепочек биополимеров, в которых имеют место перекрестные молекулярные сшивки, скрепляющие ткани, что «делает их инертными и защищает от химических атак», в частности, за счет потери растворимости. В живых тканях подобные сшивки объясняют потерю кожей эластичности с возрастом [135, 136].
Почему-то в обоих источниках [135, 136] железо названо тяжелым металлом, что не соответствует действительности (тяжелые металлы — это медь, свинец, ртуть, кадмий и пр.). Вероятно, доктор М. Швейцер на презентации сказала: «железо и тяжелые металлы», поскольку свободные радикалы, действительно, генерируются тяжелыми металлами тоже [137]. Ну, а авторы [135, 136] несколько перепутали, поскольку железо — оно, конечно, по жизни тяжелое. Странно, что перепутали даже в бюллетене университета Северной Каролины [136]. (Нам не хочется и думать, что доктор М. Швейцер и профессор Дж. Хорнер, придумывая свою гипотезу со свободными радикалами, при этом были не способны отличить железо от тяжелых металлов.)
Сама же изложенная идея для того, кто как-то занимался исследованиями биологических эффектов оксидантов и антиоксидантов, представляется странной. В состав гема гемоглобина входит двухвалентное железо. И просто удивительно, что катализируемой ионами двухвалентного железа известной реакции Фентона, когда образуется крайне активный гидроксильный радикал (все разрушает), приписывается способность стабилизировать биополимеры через внутренние перекрестные сшивки. Реакции с участием ионов железа хорошо известны, поэтому научных источников много. Только для порядка даем ряд очень конкретных ссылок [138–141].
Индуцированные ионами железа свободные радикалы агрессивны, они вызывают распад биополимеров (белков, липидов, углеводов, нуклеиновых кислот) в конечном счете даже до очень низкомолекулярных компонентов [138–141]. У М. Швейцер и Дж. Хорнера имеется смешение понятий об эффектах свободных радикалов. Действительно, для органических полимеров (полиэтилен) реакция полимеризации в промышленности инициируется свободными радикалами. Какое это имеет отношение к эффекту индуцированных ионами двухвалентного железа активных радикалов (преимущественно гидроксильного) на уже сформированные биополимеры?
В белках кожи с возрастом, в самом деле, могут образовываться за счет окисления внутримолекулярные сшивки, но из этого не следует, что молекулы с становятся неизмеримо стабильнее к действию микробов, протеолитических ферментов после смерти и пр. Попробуйте обработать белки кожи ионами железа — вряд ли при этом образуются стабильные полимеры со сшивками. И т.д., и т.п.
Причем как-то забывается, что перечисленные выше в табл. 2 и 3 остатки белков сохранились в сухом, практически дегидратированном виде (а иначе бы не сохранились), поэтому если и протекали свободно-радикальные реакции в костях, то только в течение короткого периода после их захоронения, пока все не высохло.
Таким образом, рассмотренная наукообразная гипотеза [135, 136] применительно к сохранению гемоглобина в кости тираннозавра [55] не выдерживает критики и, к тому же, не объясняет консервации других белков, что рядом с железом «не лежали» (гликопротеинов моллюска, например).
Вот и все гипотетические механизмы сохранности нестойких полипептидных структур, которые были предложены до настоящего времени (вернее, ничего более нам не встретилось).
4.4. Сохранность белков в археологических, палеоантропологических и
других остатках «малого» возраста
По идее, если фрагменты белков, порой значительной молекулярной массы и интактности (см. табл. 2 и 3), сохраняются вплоть до «сотни миллионов лет», то любому непредвзятому человеку, стоящему на позиции здравого смысла, кажется ясным, что в останках исторических и доисторических людей, которые «моложе» на три — четыре порядка, белки должны, как правило, детектироваться несравнимо лучше. Но, как и в случае с сохранностью стабильных и лабильных белков, мы и здесь не наблюдаем столь уж явных отличий. Ранее, в [1], мы представляли соответствующую сводку данных; за прошедший период нам стали известны и некоторые другие факты.
Альбумин. Радиоиммунологическими и иммунохимическими методами этот белок идентифицирован в останках мамонтов [37, 38, 142], мастодонта [40], ископаемого бизона, неандертальца и Homo erectus [37, 38]. В то же время, хотя альбумин был зафиксирован во всех исследованных в [143–145] костях исторических и доисторических людей от XVII в. до бронзового века (максимум 4200 лет назад), качество детекции очень сильно зависело от древности материала. Сходным образом, альбумин в костях перуанцев из захоронений до открытия Америки оказался весьма сильно измененным в процессах диагенеза [146]. В работе [147] указывается, что в археологических образцах белки крови определяются плохо, неоднозначно.
Тем не менее, мы можем вспомнить (см. табл. 3), что данный белок идентифицировался в останках обезьяны и гоминид, которым приписано «около двух и даже до восьми миллионов лет». Разница в возрасте между останками бронзового века, в которых альбумин уже определялся не очень-то хорошо, и костями обезьяны в «восемь миллионов лет» составляет 2000 раз...
Иммуноглобулин G. В рамках судебной медицины показано, что иммуноглобулин G сохраняется в пятнах крови на ткани, закопанной в почву, не более 4–8 месяцев [148]. И, в то же время, этот белок зафиксирован в останках доисторических детей [149] и в костях гоминида и лошади возрастом в «1,6 млн. лет» [108, 122]. В последнем случае можно было дифференцированно определить иммунохимически белки гоминида и лошади (иммунного перекреста не наблюдалось) [122]. Столь высокая сохранность для очень лабильного белка кажется просто удивительной, а вот судебно-медицинские исследования [148] таковыми не кажутся.
Гемоглобин. Не кажутся удивительными и выводы из исследований гемоглобина иммунохимическими методами в костях возрастом от 15-ти до 3000 лет: в наиболее старых образцах белка было мало [150]. Сходные результаты получены и при изучении останков людей от Древнего Рима до энеолита (максимум 5–7 тыс. лет назад) [41, 151]. Но давайте вспомним иммуногенные фрагменты гемоглобина тираннозавра возрастом в «65 млн. лет» [55]. Как же он сохранился-то, даже входя в состав неких гипотетических «кристаллов», ведь разница в сроках с энеолитом составляет 10 тыс. раз?
Остеокальцин. Здесь, как кажется поначалу, особых сюрпризов мы не наблюдаем: белок недавно выявили в костяках двух «неандертальцев», и не только выявили, но даже определили значительную часть аминокислотной последовательности [152]. Это, вроде бы, подтверждает наше сделанное выше заключение, что остеокальцин сохраняется максимальное время. И все бы хорошо, если бы именно с остеокальцином не обнаружились особые странности, которым мы уделили ниже специальный подраздел.
Коллаген. По идее, если вспомнить коллагены возрастами в «десятки миллионов лет», а также обилие этого белка (максимален в организме [99]), то уж для останков людей должно было накопиться видимо-невидимо уверенных определений коллагена. И в самом деле: для скелетов и замороженных мумий человека возрастом от 0,55 до 12 тыс. лет [111, 153–158], а также для мамонтов и бизонов [37, 38, 159], коллаген детектирован. Но уже для хряща с оцененным возрастом в «20–25 тыс. лет» белок был сильно деградирован [160], а в останках ряда «неандертальцев», обретенных в различных районах мiра (Ирак и Хорватия), найти коллаген не удалось [152]. Общим для этих останков «неандертальцев» является их древний оцененный возраст — «40 и 75 тыс. лет» [152]. Но из табл. 2 мы видим, что коллаген прекрасно обнаруживается в костях, насчитывающих по крайней мере «десятки миллионов лет»... Разница в возрасте с хрящом [160] — 2–3 тыс. раз.
Все сказанное относительно отсутствия качественных отличий между определениями в археологическом и прочем материале по сравнению с палеонтологическим приходило в голову не только автору представленного вам труда. Так, относительно нахождения в останках гоминид и лошади испанскими исследователями в 1998 и 2002 гг. альбумина и иммуноглобулина G возрастами в «1,6 млн. лет» [108, 122] некто П. Палмквист (P. Palmqvist) отмечал следующее (то ли в рецензии на [108] (см. в [122]), то ли в отдельной статье [161]): имеются расхождения между данными испанских авторов и цитированными нами выше исследованиями из области судебной медицины, где показана плохая сохранность белков уже через несколько месяцев после захоронения [145, 148]. На эту инсинуацию П. Палмквиста последовал распространенный ныне ответ, что обычно белки сохраняются, действительно, плохо, но если они попадут в специальные условия, то ничто не помешает им выдержать не месяцы, а «миллионы лет» [122]. Нравится кому такое объяснение, или же нет, пусть каждый решает сам. На мой же взгляд оно не способно объяснить продление сроков жизни не просто в какие-то там десять раз или несколько больше, а на целых три — четыре порядка. И не способно объяснить довольно большое число успешных определений белков в самых различных образцах геологического порядка времени (табл. 2 и 3). Неужели все они попадали в столь чудесные условия? (До радиации же, которая все «сотрет», мы еще не дошли.)
Конечно, если меня спросить с пристрастием: «Способны ли выдержать белки миллионы и миллиарды лет в принципе?», то я отвечу: «В принципе, способны: в толстом свинцовом контейнере без ионизирующей и УФ радиации, при температуре близкой к абсолютному нулю по Кельвину, в вакууме, в темноте, в стерильных условиях и без воды». Но в естественной среде подобный комплекс факторов вряд ли мог сформироваться.
4.5. Остеокальцин: «чехарда» сроков сохранности
Ведущими исследователями остеокальцина в ископаемых останках ныне являются Кристина Нильсен-Марш (Christina Nielsen-Marsh) из университета в Ньюкастле, Великобритания, и Пеги Эстрем (Peggy Ostrom) из Мичиганского государственного университета. Ранее в своих работах они идентифицировали остеокальцин в останках ископаемых бизонов возрастом «до 300 тыс. лет» [98], а в настоящее время их рекордом является остеокальцин из костей мускусного быка возрастом в «500 тыс. лет» [135]. В работе [1] мы провели соответствующие расчеты по первичному материалу [98] и выполнили следующий график (рисунок 5).
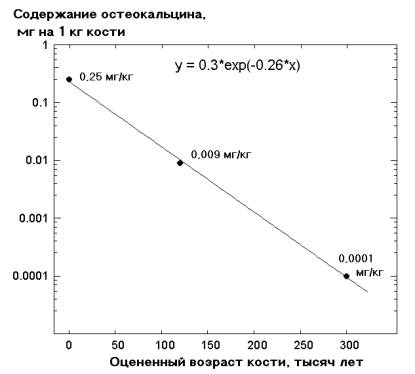
Рисунок 5. Степень сохранности остеокальцина в костях древних
бизонов в зависимости от возраста останков [1] (определение
радиоиммунологическим анализом с антителами к остеокальцину коровы [98]).
Из графика на рис. 5 видно, что содержание остеокальцина снижается в ископаемых костях четко по экспоненциальной зависимости. Это и неудивительно — такая же закономерность характерна для многих физико-химических и биохимических процессов распада. (Но наш расчет подтверждает действительную реальность стандартной зависимости для конкретного белка — остеокальцина.) Еще из графика следует, что содержание остеокальцина уже в останках возрастом в «300 тыс. лет» крайне мало, в килограммах кости определяются прямо-таки следовые количества белка. Однако в последние годы он был идентифицирован К. Нильсен-Марш и П. Эстрем в костях возрастом уже в «полмиллиона лет» [135]. Что ж, это не очень удивляет: возможно, здесь сказываются вариации в степени сохранности костей в различных местах и условиях. Но десятилетием ранее остеокальцин был четко идентифицирован иммунохимическими методами в останках, возраст которых просто несоизмерим с показателем для костей бизонов [98] и мускусного быка [135]. Частично уже приводившиеся выше данные сведены в таблицу 4.
Таблица 4. Определение остеокальцина в ископаемых костях
|
Исследованные
кости |
Оцененный
возраст |
Ссылка |
|
Два иракских «неандертальца» |
75 тыс. лет |
[152] |
|
Бизоны |
120 тыс. лет;
первоначальный уровень* снижен в 28 раз |
[98] (расчеты см. в [1]) |
|
300 тыс. лет;
первоначальный уровень* снижен в 2500 раз; содержание очень мало — 0,0001 мг на 1 кг кости |
||
|
Мускусный бык |
500 тыс. лет |
[135] |
|
Бык (кости), грызун (зубы) |
До 13 и 30 млн. лет |
[39] |
|
Тираннозавр |
65–67 млн. лет |
[32, 113] |
|
Четыре динозавра Юрского и
Мелового периодов |
73–75 и 150 млн. лет |
[118–120] |
|
Игуанодон |
120 млн. лет |
[121] |
*
По сравнению с содержанием остеокальцина в костях современной коровы [98].
Если считать, что в костях возрастом 300–500 тыс. лет иммунохимическими методами определяются следовые количества остеокальцина, то каким образом он почти «систематически» выявляется сходными методами в различных останках динозавров (пока шести), обнаруженных в различных регионах? Оцененный же возраст этих останков динозавров больше, чем у образцов бизонов и мускусного быка в 130–400 раз.
И мы не можем сбрасывать со счетов, что для уверенной идентификации остеокальцина иммунохимическими методами от исходной молекулы белка должно оставаться ни много ни мало, а не менее 12–16% (расчет см. в подразделе 4.1). То есть в костях динозавров юрского и мелового периодов, а также игуанодона (все — от «73-х до 150 млн. лет» — см. табл. 2 и 4), молекула остеокальцина уменьшилась всего в 6–8 раз. Получается нечто вроде «периода полураспада» (молекулы по размеру), величина которого для «8-ми раз» составляет от «24-х до 50-ти млн. лет». Это, знаете, впечатляет: полураспад, сходный с параметром элемента кюрия — 247Cm (15,8 млн. лет), открытого четой Кюри. Берем молекулу белка остеокальцина, кладем и уходим. Возвращаемся через 24–50 млн. лет, и имеем нетронутую половину от исходной полипептидной цепи. Хорошо бы так было с процессом денежной инфляции.
Словом, все получается очень странно, как и с археологическими и палеоантропологическими образцами. Но странно только в том случае, если мы не признаем, что никаких «миллионов лет» не было, и что возраст всех ископаемых остатков с определяемым остеокальцином более или менее равновелик.
Если находиться на креационных позициях, то все встает на свои места.
Вопрос же о воздействии на биомолекулы радиации в течение «десятков миллионов лет», и во что это должно было вылиться, мы рассмотрим ниже.
5. КЛЕТКИ И МЯГКИЕ ТКАНИ КАК ТАКОВЫЕ В ОСТАНКАХ ВОЗРАСТОМ В «ДЕСЯТКИ
МИЛЛИОНОВ ЛЕТ»
5.1. Реальность опять невероятней фантастики
Если бы кто-нибудь показал специалистам подобное название главы лет пять — десять назад, то реакции могли быть малопредсказуемыми. В частности, большинство просто крутили бы пальцем у виска. И если бы мне показали такое хотя бы около трех лет назад, когда я готовил первый обзор по молекулярной палеонтологи [1], то и я бы не поверил, несмотря на идентификацию польскими авторами под микроскопом структур, похожих на сосуды и клетки, в кости динозавра еще в 1966 г. [19–21, 24–29, 31]. Их косвенные данные были подтверждены напрямую в 1990-х гг., да и то со скандалом и замалчиванием. И только в 2005–2006 гг. отпали последние сомнения.
Пока не наступил тот 2005 г., я, к примеру, писал в своем обзоре [1] (который готовил в 2004 г.) о польских результатах следующее:
«Про видимые под микроскопом структуры, очень похожие на остеоциты и эритроциты, могут сказать, что это артефакты, обусловленные внешними воздействиями на кость в течение длительного времени. Даже то, что в районе эритроцитов обнаружено намного большее содержание железа.., не является окончательным доказательством: всегда можно предположить, что некие железосодержащие бактерии почему-то облюбовали именно эти места в какой-то момент из многих миллионов лет».
А вот как я изволил рассуждать в [1] про упоминание эритроцитов тираннозавра доктором М. Швейцер с соавтором в научно-популярной статье 1997 г. [44]:
«И закончим про кровь тираннозавра. Понятно, что прямого доказательства, являются ли именно эритроцитами столь похожие на них под микроскопом образования, получить нельзя. Не спросишь ведь их под микроскопом: «Ты эритроцит или нет?». Да и мертвые они уже были... не написано на тех кружочках, что они именно красными кровяными клетками являются. Всякие морфологически подобные артефакты за долгое время накопится могут. Хотя, возможно, под электронным микроскопом и можно специфически именно эритроциты идентифицировать, однако я этого не знаю, не специалист тут...».
А ведь я давно стою на
креационных позициях... Что уж тут говорить о тех, кто осуществлял свой анализ
фактов с позиций эволюционных...
Но время, даже весьма короткое, часто расставляет все на свои места. То, что казалось невероятным, становится обыденностью. Рубежом, так сказать, «новой эры» явился 2005 г., когда доктор М. Швейцер с соавторами наконец-то описали в авторитетном международном журнале «Science» свое открытие. В костях ряда динозавров после удаления минеральной составляющей идентифицировались гибкие, прозрачные сосуды, содержащие внутри красные эритроциты с отчетливыми ядрами, а также клетки кости — остеоциты со всеми их особенностями. И фибриллы костного матрикса (органического сетчатого образования) [32, 113].
Теперь, с 2005 г., в мiре молекулярно-клеточной палеонтологии мы живем в другое время. В этом «новом времени», чтобы оставаться на позициях геологических возрастов ископаемых останков и эволюционизма, приходится попирать прежние представления, факты и логику физической химии, радиационной биофизики, биохимии, гистологии и клеточной биологии.
5.2. Первые относительно известные факты и их интерпретация
В начале 1990-х гг., при исследовании «недоокаменевшей» кости тираннозавра [1], доктор Мэри Швейцер с сотрудниками (США) обнаружили под микроскопом эритроциты:
«Лаборатория наполнилась возгласами изумления, когда я сосредоточила [внимание] кое на чем внутри сосудов; на том, чего ни один из нас никогда не видел когда-либо ранее: крошечные круглые объекты, прозрачные, красные, с темным центром. Когда коллега бросил всего один взгляд на них, то он закричал: «Вы обнаружили эритроциты! Вы обнаружили эритроциты!» (цитировано по [162]; здесь и далее перевод цитат мой. — А.Л.).
Срез кости под микроскопом один из сотрудников показал патологоанатому, и тот тоже сказал: «Вы понимаете, что нашли эритроциты?» [163].
Никто не мог во все это поверить, а про работу польских авторов из далекого 1966 г. [19] и не вспомнили. (Да и вряд ли они хорошо помнили, где та Польша находится.) Когда доктор М. Швейцер показала слайд с эритроцитами тираннозавра своему шефу, профессору Джеку Хорнеру, то тот не нашел ничего лучшего, чем предложить ей для докторской диссертации (Ph.D.) доказать факт, что видимые под микроскопом эритроциты на самом деле таковыми не являются, что это просто диагенетические артефакты [163]. Но в процессе выполнения темы докторантом было доказано, как мы знаем [2, 3], обратное. В 1997 г. доктор Мэри Швейцер вместе с соавтором опубликовала в научно-популярном американском журнале «Earth» статью под названием «Реальный Парк Юрского периода», где было фото эритроцитов с подписью: «Кровь из камня» [44]. Журнал тот вскоре прекратил свое существование, но статья наделала много шуму. Фрагменты из нее цитировались известным креационистом, доктором Карлом Виландом в 1997–2002 гг. [162, 164, 165], из находки делались далеко идущие выводы, которые, затем, подвергались обсуждению и соответствующей критике эволюционистом доктором (Ph.D.) Джерри Хардом (G.S. Hurd).
Этот доктор Дж. Хард на сайте «The Talk Origins Archive. Exploring the Creation/Evolution Controversy» [166] выступает с постоянной критикой позиции креационистов в области молекулярно-клеточной палеонтологии, однако в конечном счете все время попадает пальцем в небо, поскольку последующие открытия опровергают его прежние критические выводы относительно правоты сторонников «молодой Земли».
После публикации в 1997 г. в «Earth» указанных «Парка Юрского периода» и «Крови из камня» в научном и околонаучном мiре эволюционистов поднялся шум, доктору М. Швейцер досталось за «рекламность» и за то, что она представляет репортерам результаты своих еще незаконченных работ «в преувеличенном и спекулятивном виде» [163, 166]. Позднее, в 2006 г., доктор М. Швейцер так вспоминала этот момент: «Я раздразнила гусей (goose bumps), потому что каждый знает, что подобные вещи не сохраняются в течение 65 млн. лет» [163] (см. также в [167]). Возникли трудности с публикациями в научных журналах статей по исследованию кости тираннозавра [163]. А шефу доктора М. Швейцер, профессору Дж. Хорнеру, пришлось неоднократно оправдываться, говоря нечто вроде: «Да не нашли мы там на самом деле никаких клеток крови!» [166]. И он делает это до сих пор, несмотря на то, что в 2005 г. в «Science» были опубликованы однозначные данные по сосудам, эритроцитам и остеоцитам в костях четырех динозавров. Например, в интервью 2006 г. «National Geographic News» профессор Дж. Хорнер сказал насчет полностью сохранившихся биоструктур следующее: «Мы не знаем, из чего они состоят. Они выглядят как кровеносные сосуды. Они эластичны и во всем подобны им, но состоят ли они из исходного вещества или нет, мы не знаем» [168]. Похоже, профессор ожидает найти такие минералы, которые, заместив в сосудах органику, оставят их «гибкими и прозрачными». И почему-то забывает, что более 50% элементов в образцах составлял углерод (ниже).
Эволюционист Дж. Хард в 2004 г. [166] пенял креационисту доктору К. Виланду на то, что тот когда-то поверил статье в научно-популярном журнале «Earth» и постоянно яро популяризует эритроциты тираннозавра: «Применительно к «красным кровяным клеткам» динозавра, аргумент, агрессивно проталкиваемый доктором К. Виландом, является активным и поразительным опровержением [действительного] факта». Далее сказано, что нечего было опираться на «неокончательные» данные в научно-популярном журнале, и что сами авторы той статьи 1997 г. писали, что де, это у них не эритроциты как таковые, а «в лучшем случае производные [клеток] крови, модифицированные тысячелетиями геологических процессов» [44, 166]. (Почему-то написали для костей тираннозавра «тысячелетия» — «millennia», а не «millions», тем более не «tens millions» — десятки миллионов [166].)
Словом, доктор Дж. Хард за год до опубликования микрофотографий окрашенных эритроцитов, остеоцитов и гибких прозрачных сосудов [32, 113] не только не верил в реальность всего такого, но и критиковал креациониста доктора К. Виланда за «ненаучность» и «спекулятивность».
Применительно же к идентификации остеокальцина в костях четырех динозавров [118] и гемоглобина в кости тираннозавра [55] Дж. Хард вновь пенял креационистам на увлечение сенсационным, и у него были слова, что никаких целых белков и эритроцитов в костях тираннозавра на самом деле не обнаружили [169].
Подобные фразы насчет отсутствия клеток динозавров как таковых имели место до того момента, пока не вышли окончательные публикации. Но даже и сейчас мы встречаем скептицизм, который частично рассмотрели в [2]. Например, в современной англоязычной энциклопедии динозавров online [170] еще в 2006 г. было сказано (выделено мною):
«Возможное открытие мягких тканей в окаменелостях динозавра. ...Исследование под микроскопом показало, что в предполагаемых мягких тканях динозавра остаются микроструктуры даже на клеточном уровне. Не ясно, из чего фактически составлен этот гибкий материал, хотя многие сообщения немедленно связали его с кино «Парк Юрского периода». Вопросы об интерпретации полученных данных и относительной важности открытия доктора Швейцер все еще не решены».
Однако время идет, подтверждений кажущейся фантастике прибавляется, и далее мы подробно рассмотрим конкретные данные.
5.3. Публикации в «Science» 2005 г.:
гибкие, прозрачные сосуды, эритроциты и остеоциты в костях четырех динозавров.
А к 2007 г. — в костях еще нескольких
Мы уже разбирали эти публикации (вернее, одну с сопутствующим материалом online) [32, 113] в своем обзоре [2]. Здесь повторяем вкратце.
Исследованные объекты. В 2003 г. в мягком песчанике в отложениях горного массива в штате Монтана был обнаружен скелет Tyrannosaurus Rex. Он получил музейный индекс MOR 1125 (аббревиатура начальных букв названия «Музей экспонатов Скалистых гор» — «Museum of the Rockies specimen») [32]. Исследование этого образца изложено в основной части работы [32], а в приложении к ней online [113] помещены в том числе результаты изучения хорошо сохранившихся костей еще трех динозавров: тираннозавров под индексами FMNH-PR-2081 и MOR 555, а также гадрозавра MOR 794 (Brachylosaurus; утконосый динозавр позднего мелового периода). MOR 555 — это экземпляр, обнаруженный в 1990 г. в том же месте, что и MOR 1125. Он был исследован еще в 1997 г. [44, 55].
Методика. Фрагменты кости деминерализовали, помещая их на семь дней в концентрированный — 0,5 моль/л (около 19%) — раствор этилендиаминтетрауксусной кислоты (ЭДТА), pH 8,0 [11]. Раствор меняли ежедневно. ЭДТА представляет собой хелатирующий реагент, который связывает кальций и магний, удаляя, таким образом, из кости неорганическую составляющую. Заметим, что это не «кислотный раствор», как интерпретируется иной раз [171], просто реактив — ЭДТА — это кислота, но используют ее динатриевую соль в слабощелочных условиях.
Материал, полученный после деминерализации, промывали, фиксировали и затем микроскопировали под световым и электронным микроскопами для идентификации микроструктур. В некоторых случаях препараты окрашивали красителем, специфичным для клеточных ядер [113].
Была также проведена экстракция белкового (пептидного) материала из кости тираннозавра MOR 1125. На стандартной методике приготовления экстракта мы останавливаться не будем, скажем только, что его использовали для детекции фрагментов белков с помощью коммерческих (т.е. тех, которые изготовляют фирмы) препаратов антител к коллагену цыпленка и к остеокальцину быка. Реакцию проводили путем иммуноферментного анализа (ELISA). Авторами были соблюдены все приходящие на ум контроли: помимо стандартных контролей, исследовали взаимодействие антител и с экстрактом из окружающей кость породы, и с экстрактом из растительного материала, ассоциированного с образцом [113].
Для деминерализованных костных препаратов проводили анализ составляющих их химических элементов. В качестве образца сравнения использовали кости и биологические структуры, полученные от современных нам страуса и цыпленка [113].
Деминерализация образцов и химический состав. В таблице 5 представлен химический состав деминерализованных препаратов сосудов из костей тираннозавра и современного страуса.
Таблица 5. Химический состав деминерализованных препаратов сосудов из костей тираннозавра MOR1125 и страуса (% от суммарного атомного состава) [113]
|
Элемент |
Атом%, MOR 1125 |
Атом%, страус |
|
C |
55,66 |
53,29 |
|
Ca |
Не идентифицирован |
0,29 |
|
Ti |
0,69 |
0,47 |
|
O |
21,18 |
38,88 |
|
Fe |
2,86 |
Не идентифицирован |
|
F |
1,95 |
Не идентифицирован |
|
Zn |
0,81 |
0,65 |
|
Na |
1,56 |
0 |
|
Al |
0,7 |
0,19 |
|
Au |
1,57 |
0,88 |
|
Si |
9,26 |
4,12 |
|
P |
0,77 |
Не идентифицирован |
|
Cl |
1,19 |
0,11 |
|
K |
1,8 |
1,12 |
Из табл. 5 видно, что для обоих образцов углерод составляет более половины материала, и, вероятно, препараты содержат преимущественно органику. Есть и кремний, но он есть и в образце от страуса, хотя, конечно, в меньшем количестве, чем для тираннозавра. Но мы все-таки не можем сбрасывать со счетов, что последний какое-то время покоился в недрах горной породы. Кальций из кости ящера удалился при деминерализации несколько лучше, чем из кости страуса, зато железо и фтор — наоборот. В целом же мы можем сделать тот же вывод, что и сами исследователи [113]: состав препаратов тираннозавра и страуса очень похож.
Недавно, в феврале 2007 г., один канд. хим. наук прислал мне следующее замечание-вопрос. Он поинтересовался, почему среди минеральных составляющих, которые, по идее, происходят из белковой органики, нет серы и азота. Отсюда следовал его вывод, что препарат деминерализовался не полностью, и что он состоял преимущественно не из органического материала, а из неорганического углерода и прочих минералов. Остатки белков если там и были, то только в столь следовом количестве, что их сера и азот просто не определились.
Но приславший замечание просмотрел, что точно такой же состав, без серы и азота, был получен (представлен) авторами и для обработанного коллагеназой (освободила клетки и сосуды от сетей костного матрикса) препарата из кости современного страуса (см. табл. 5). Последняя, согласно [113], «хранилась несколько лет при –20°C». Вероятно, такова была методика анализа химических элементов, использованная авторами, что азот и сера не определялись [32, 113]. Адекватна она или нет, мы судить не станем, скажем только, что палеонтологи и геофизики из университетов Монтаны и Северной Каролины, а также из Института Карнеги в Вашингтоне [32, 113], вряд ли могли допустить столь серьезную ошибку, «повесив» ее затем online. Главное же в их анализе — очень близкие величины для препаратов тираннозавра (не менее «65 млн. лет») и страуса (несколько лет, да еще при –20°C).
Сосуды. При исследовании деминерализованных образцов из трех тираннозавров под микроскопом выявилась разветвленная сосудистая сеть: прозрачные, гибкие, полые кровеносные сосуды. Эти сосуды проявляли упругость при манипуляциях. При повторяющихся циклах дегидратации — регидратации они все еще оставались эластичными. Был сделан вывод, что «мягкие ткани динозавра могут сохранять некоторую первоначальную гибкость, эластичность и упругость» [32].
Сохранность для различных образцов, конечно, варьировала; отдельные сосудистые структуры имели определенную степень «закристаллизованности». Но прозрачные гибкие сосуды выявлялись у всех трех тираннозавров (см. ниже рисунок 6.1, а, б). В то же время, в образцах из кости гадрозавра сосуды сохранились плохо: они были «фрагментарны и сильно изменены». Забегая вперед скажем, что, тем не менее, именно у гадрозавра почему-то лучше сохранились остеоциты по сравнению с тираннозаврами [113].
Сосуды, сходные по диаметру, строению и виду, были получены из современной кости страуса при ее деминерализации коллагеназой, удаляющей плотный волокнистый коллагеновый костный матрикс [32, 113].
Эритроциты. Внутри видимых сосудов имели место включения, которые при увеличении разрешения оказались эритроцитами. (Напомним, что у земноводных, пресмыкающихся и птиц, в отличие от млекопитающих, зрелые эритроциты имеют ядра.) Для этих образований имело место характерное окрашивание (от темнокрасного до темнокоричневого) и темные ядерно-подобные включения (рис. 6.1 в, г). Вновь картина была полностью аналогичной виду эритроцитов страуса [32, 113]. Позднее доктор М. Швейцер в своих выступлениях показывала одновременно две микрофотографии образцов, приговаривая: «Одним из этих клеток 65 млн. лет, а другим — 9 месяцев. Можете вы сказать мне, какие из них какие?» [135].
Отчетливые эритроциты к 2005 г. были выявлены в образцах костной ткани трех тираннозавров. У гадрозавра же эритроциты по сравнению с его остеоцитами производили худшее впечатление [113].
Остеоциты. В костях всех четырех динозавров в [32, 113] обнаружены остеоциты, да не просто — а в ряде случаев с ядрами и с филоподиями-«ножками» (см. рис. 6.1, д, е). У MOR 1125 они могли выходить из ткани в раствор, в то время как для других образцов такого не наблюдалось.
Но доктор Р. Павлички из Польши снова оказался впереди: он описал остеоциты в костях динозавра еще в 1975 г. [21].
Фотодокументы. На рисунке 6.1 нами приведены избранные фотографии микроструктур из деминерализованных костей различных динозавров [113]. Всего же в [32, 113] опубликовали массу микрофотографий. Ранее мы не осмеливались представить большинство из них за недостатком места [2], но здесь, в обзоре, имеем подобную роскошь. На рис. 6.2–6.4 приведены фотодокументы так, как они опубликованы в оригиналах — печатной статье в журнале «Science» [32] и приложении к ней online [113].
Мы специально даем материал аутентично оригиналам, даже без перевода. «Это документ, между прочим»[16]. Пусть каждый, взяв англо-русский словарь, убедится в том, что в костях тираннозавров (на фотоиллюстрациях приведены результаты исследования нескольких) имелись гибкие органические структуры, отвечающие на манипуляции сжатием — расжатием, обнаружено множество эритроцитов внутри прозрачных сосудов, другие клетки... прочее... Что говорить — надо смотреть, поэтому мы и приводим здесь почти всё (нет только одного малого непринципиального фото остеоцита с филоподиями). По нашему мнению, выдержать зрелище, отображенное ниже на рис. 6.1–6.4, сможет, думается, далеко не каждый, а только самый упертый сторонник длительных геологических эпох, обеспечивавших, согласно предрассудкам, базу для макроэволюции.
Рисунок 6.1. Сосудистые и ядерные микроструктуры из
деминерализованных костей динозавров
Гибкие,
прозрачные, разветвленные сосуды (а, б); фрагмент сосуда в прозрачном упругом
костном матриксе (в); содержимое
этого сосуда — эритроциты с ядрами (г); остеоциты в матриксе (д); изолированный остеоцит со специфическими
структурами (е) [113].
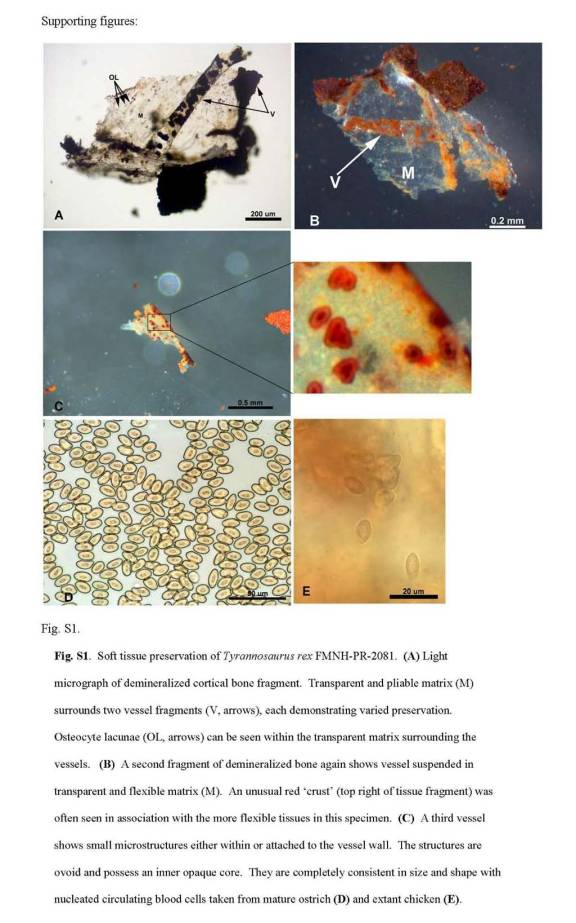
Рисунок 6.2. Оригинальный фотоматериал из [113] по исследованию мягких тканей, клеток и биоструктур в кости тираннозавра FMNH-PR-2081.
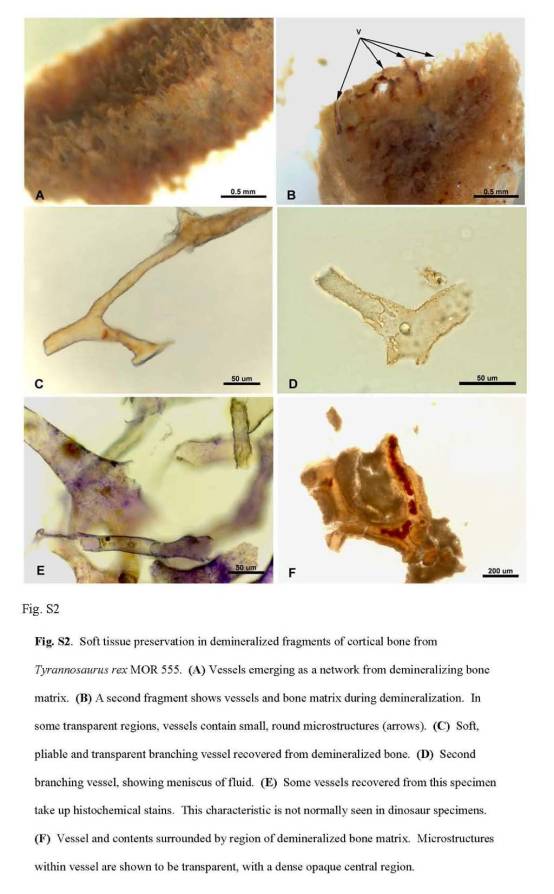
Рисунок 6.3. Оригинальный фотоматериал из [113] по исследованию мягких тканей, клеток и биоструктур в кости тираннозавра MOR-555.

Рисунок 6.4. Оригинальный фотоматериал из [32] по исследованию мягких тканей, клеток и биоструктур в костях тираннозавров.
Фрагменты белков в костях тираннозавра. В данном случае был исследован один образец — MOR 1125. Для препаратов из костной ткани этого тираннозавра, из окружающей ее породы и ассоциированных с ней растительных остатков, а также для образцов из костей цыпленка и страуса, проводили реакции с антителами к коллагену цыпленка и к остеокальцину быка. Обнаружено, что контрольные образцы (порода и растения) не связывались с антителами. Препарат же тираннозавра давал очень отчетливые реакции в обоих случаях, хотя, конечно, более слабые, чем для образцов современных нам цыпленка и страуса [113].
Таким образом, в кости тираннозавра имелись не распавшиеся до конца фрагменты и коллагена, и остеокальцина, что, конечно, в свете данных, приведенных выше в табл. 2, не кажется удивительным.
Дальнейшее подтверждение находок 2005 г. В 2006 г. доктор М. Швейцер с соавторами продолжала исследовать кости ископаемых животных. Согласно [135], эксперименты с деминерализацией были повторены с более чем дюжиной других образцов динозавров и прочих ископаемых животных. Сказано, что для приблизительно половины препаратов «получены замечательно последовательные результаты», и что вид под микроскопом «был неотличим от образцов современных тканей» [135]. Рекордом, видимо, надо пока считать биоструктуры из костей гадрозавра возрастом в «80 млн. лет» [136]. А там, глядишь, и до эритроцитов какого-нибудь игуанодона в «120 млн. лет» доберутся.
Нами было опубликовано
обсуждение исходных результатов доктора М. Швейцер по четырем динозаврам в
2006 г. [2], а последующая «полудюжина» находок вплоть до «80 млн.
лет» упомянута в обзоре начала 2007 г. [3]. И вот только что была
обнаружена новая аналогичная публикация доктора М. Швейцер с соавторами,
статья за январь 2007 г. [172]. В ней однозначно подтверждается реальность
всего выявленного ранее в 2005 г. [32, 113] для костей тираннозавра:
гибкого и волокнистого костного матрикса, гибких,
полых, прозрачных сосудов,
интраваскулярного (внутрисосудистого) материала, включая структуры, морфологически
напоминающие эритроциты позвоночных, а также остеоцитов с внутриклеточным
содержимым и с гибкими филоподиями.
Авторы [172] отмечают, что минерализация костей может быть неполной[17],
и что мягкие ткани, де, способны сохраняться лучше, чем считалось ранее. Ни в
каких «миллионах лет» авторы [32, 113, 172] не сомневаются — промывка,
вернее наоборот, засорение мозгов капитальное. И, тем не менее, доктор
М. Швейцер со своими соавторами в 2007 г. договорилась до того, что
остеоциты динозавров с сохранившимися ядрами «могут быть источником
генетической информации» [172]. В такое не верим даже мы, разве что речь идет
об «информации» по типу «ДНК-неандертальца» [12, 13]. когда желаемое принимают
за действительное (обоснования см. в [1] и, соответствующий краткий
очерк — в [5])».
Основной механизм странной и систематической сохранности мягких тканей в костях динозавров, согласно последней гипотезе доктора М. Швейцер, заключается в образовании резистентных (устойчивых) полимерных биоструктур за счет формирования внутри- и межмолекулярных сшивок, индуцируемых в результате атак свободных радикалов, генерируемых ионами двухвалентного железа, освобождающегося из распавшегося гемоглобина [135, 136] (это я тут по-научному, используя термины, принятые в области изучения окислительных процессов, изложил). Несостоятельность данной гипотезы мы уже рассмотрели выше в подразделе 4.1, пункт 5. Более ничего вразумительного, кроме, опять же, формирования «внутренних кристаллов», в оригинальных публикациях М. Швейцер с соавторами [32, 172] мы не находим.
Так что, господа, в 2006–2007 гг. данные о мягких тканях, окрашенных эритроцитах и остеоцитах (см. рис. 6.1–6.4) не только не опровергнуты, но даже прибавились. Все становится, так сказать «обыденностью»: «Множество окаменелостей динозавров могут иметь внутри мягкую ткань» (доктор М. Швейцер). Этой молодецкой фразой главного молекулярно-клеточного палеонтолога назвали в 2006 г. статью в «National Geographic News» [135].
Значит, теперь «множество окаменелостей могут...». Но совсем недавно, считанные годы назад, обнаружив под микроскопом сосуды динозавра, доктор М. Швейцер не поверила своим глазам: «Когда окаменелость распалась, то остались прозрачные сосуды. «Это был полный шок», — рассказывала Швейцер. — «Я не поверила, пока мы не повторили опыты 17 раз» [171].
Хорошо, что не «до седмижды семидесяти раз» (Мф. 18:22). А то мы так и не увидели бы публикаций...
Теперь же: подумаешь — сосуды и эритроциты динозавров... Вот у С. Спилберга и «BBC — живой мир», у тех, действительно, у тех — живые тираннозавры. А здесь — ерунда какая-то... Сосуды и эритроциты возрастом в «десятки миллионов лет» — эка невидаль... Сторонники биогенеза и эволюции еще и не в такие чудеса заставят поверить априори.
Но мы, креационисты, и здесь впереди: у нас на руках такие карты применительно к информации о «чудесах сохранения», какие никому из эволюционистов и в страшном сне присниться не могут (ниже в разделах 7 и 8).
6. КОСТНЫЙ МОЗГ ВОЗРАСТОМ В «ДЕСЯТЬ МИЛЛИОНОВ ЛЕТ»
6.1. Самая нестойкая ткань может храниться с миоцена
Прошло чуть более полутора лет с момента выхода публикаций о сосудах и клетках в костях динозавров (март 2005 г.) [11, 32, 113], а в области молекулярной палеонтологии появились результаты в чем-то еще более невероятные, чем даже исследования доктора М. Швейцер с сотрудниками. В августе 2006 г. в «Geology» (журнал Геологического общества США) были опубликованы данные группы авторов во главе с исследовательницей из Ирландского университетского колледжа в Дублине Марией Макнамарой (Maria McNamara)[18]. Эти результаты получены в рамках ее будущей диссертации (Ph.D.). В отложениях древнего озера в Испании были обнаружены остатки более чем сотни амфибий — лягушек и саламандр, причем в десятой части костей сохранился костный мозг с клетками и жировыми прослойками, визуально обычного вида [173]. Важность открытия в том, что останкам этих амфибий палеонтологами приписывается возраст в «10 млн. лет», ибо они найдены в миоценовых слоях. Данное исследование и его руководительница М. Макнамара за прошедшие месяцы уже преданы широкой гласности в ряде иноязычных научно-популярных изданий и мiровых СМИ, причем без всяких сомнений в достоверности «10 млн. лет» (см. к примеру, [168, 174, 175]).
Костный
мозг представляет собой ткань, содержащуюся в полостях костей позвоночных животных
и человека. В красном костном мозге образуются форменные элементы крови —
эритроциты, лейкоциты и тромбоциты, а желтый костный мозг состоит главным
образом из жировых клеток. Многие клетки костного мозга, являющиеся
предшественниками клеток крови, постоянно делятся; данная ткань в организме
является системой клеточного обновления [176].
Но значительный пул костного мозга занимают незрелые и зрелые эритроциты, которые, как следует из данных доктора М. Швейцер, сохраняются, де, даже «десятки миллионов лет» [11, 32, 113, 172]. В то же время известно, что трупный автолиз (самопереваривание) костного мозга начинается уже через несколько часов после смерти [176]. А в целом — каждому биологу или медику ясно, что клеточный костный мозг — ткань очень нестойкая и ранимая.
И вот, «окаменевшие кости миоценовых лягушек и саламандр содержали хорошо сохранившиеся остатки костного мозга» [168, 173–175]. В костях амфибий возрастом «10 млн. лет» идентифицирована трехмерная органическая структура с оригинальной текстурой ткани, с красной и желтой окраской отчетливо видных гемопоэтических и жировых клеток, с остеокластами (мезенхимными костными клетками) и сосудами (рисунок 7).

Рисунок
7. Костный мозг в останках миоценовой
лягушки из Испании возрастом порядка «10 млн. лет». При разных разрешениях
микроскопа видны клетки с мембранами, желтый и красный кровяные ростки и пр.
[173].
Помимо лягушек и саламандр сообщалось также, что фрагменты костного мозга были обнаружены в остатках головастика того же возраста [173, 177].
Цитаты из высказываний М. Макнамары [168, 174]:
«Костный мозг сохранился на органическом уровне. Оригинальный цвет ткани сохранился. Как и у современных лягушек, во внутренней зоне костей видна жировая ткань желтого цвета, окруженная внешней зоной красного костного мозга».
«Даже простое открытие того, что костный мозг имеет красный цвет, позволяет сказать, что эритроциты возникали в костях древних амфибий, а не в селезенке, как у современных саламандр».
Необходимо отметить, правда, что молекулярный анализ ископаемого костного мозга пока только проводится [168, 173], но ни авторы этого исследования (см. выше первую цитату), ни мы, глядя на фотографию (см. рис. 7) теперь не сомневаемся, что в подобных структурах будут обнаружены и фрагменты белков, и, видимо, липидов. Что ж тут удивительного, если ныне общепризнано, что клетки и органические структуры могут спокойно сохраняться в течение сроков, больших в 6–8 раз, а именно — в костях динозавров [11, 32, 44, 55, 113, 135, 163, 167, 171]. По сравнению с тираннозаврами мелового периода доктора М. Швейцер миоценовые амфибии М. Макнамары с их костным мозгом в каких-то «10 млн. лет» кажутся просто жалкими.
Кто-то может подумать, что древнее озеро Испании эндемично в смысле сохранения костного мозга миллионы лет, и что М. Макнамара всюду указывает на уникальность своих образцов. Но нет, исследовательница, напротив, считает, что костный мозг повсеместен для миллионолетних костей, стоит только их разломать [173, 177]. Ее спросили, почему же до этого никто не догадался глянуть внутрь ископаемых костей, которые рассматривали еще со времен профессоров Р. Оуэна и Э. Челленджера из «Затерянного мiра». Ответ прост, и заключается он в том, что внутрь не смотрели, поскольку «нечего портить вещь»:
«Было множество причин, по которым окаменевший костный мозг никогда не находили прежде. Поскольку костный мозг разлагается очень быстро, когда люди умирают, то никто никогда не думал, что он может сохраниться. Кроме того, чтобы увидеть кости изнутри, вы должны их разломать; очевидно, что если вы имеете хорошие образцы музейных окаменелостей, то вы не собираетесь получать разрешение их разломать» [177, 178].
В случае же испанских миоценовых лягушек и саламандр костей было много, доставались они трудно, и некоторые при раскопках, транспортировке и хранении поломались. Вот тогда-то, впервые за пару сотен лет, и догадались взглянуть под микроскопом на нутро ископаемых костей [173, 177, 178].
Надо, правда, заметить, что костный мозг может сохраниться только в, так сказать, «свежих» ископаемых костях. Если исследователи помещали найденные образцы в консерванты, то костный мозг в активной среде должен был немедленно распасться. Аналогично и с сосудами и клетками в костях динозавров. Когда впервые изучали на этот предмет бедро тираннозавра в 1990-х гг., то, как отмечал профессор Дж. Хорнер, этот образец был слишком велик для транспортировки вертолетом, кость поделили на части, в результате чего доктору М. Швейцер достался кусок из середины, а поскольку кость была поломана, то консерванты решили не применять и т.д. [179].
Но как же все-таки ископаемый костный мозг может сохраняться? Понятно, что авторы оригинального исследования древних амфибий [173], если они верят в свои «10 млн. лет», должны были предложить механизмы удивительной стойкости и сохранности нестойкой и плохо сохраняющейся ткани. «Механизм» Марией Макнамарой был предложен (см. ниже). И он кажется не менее чудным, чем у другой «не просто Марии» — доктора Мэри Швейцер.
6.2. Опрос патологоанатома
В 2006 г. я провел опрос нашего патологоанатома. Это специалист с очень большим стажем, профессор, заведующий лабораторией. У нас (и не только) он пользуется большим авторитетом. Я не стал ничего говорить ни о креационизме, ни о чем подобном, а просто спросил вначале, как долго может сохраняться в трупе костный мозг. Воспроизвожу диалог:
П.: «Это смотря как хранить».
Я: «Ну, наилучшим образом».
П.: «Неделю, дней десять в холодильнике сохранится».
Я: «Да нет, вы, наверное, имеете в виду живые клетки, а я спрашиваю, как долго будет сохраняться их морфология под микроскопом».
П.: «Нет, не живые — живых там не останется, просто их можно будет дифференцировать по типам, окрасить специальными красителями, увидеть под микроскопом и т.п.»
Я: «А вот у миоценовых лягушек «10 млн. лет» клетки в костном мозге сохраняются...».
Понятно, что ответом было недоумение. Тогда я послал нашему патологоанатому по электронной почте фото костного мозга миоценовой лягушки (см. выше рис. 7) и фото сосудов тираннозавра с эритроцитами и остеоцитами (рис. 6.1), предварительно уточнив, что материал — из публикаций в академических журналах США. Сомнений в подлинности фото у патологоанатома не возникло, хотя он и нашел одну «неграмотность»:
П.: «Тут неверно на фото с остеоцитами тираннозавра».
Я: «Что — не клетки, что ли? Так это не ко мне — фото из оригинала».
П.: «Нет, там написано в подписи, что представлен «остеоцит с филоподиями-ножками», но там этого нет».
Я: «А что есть?»
П.: «Это у остеоцита специальные каналы, через которые остеоциты друг с другом контактируют, но не филоподии; неграмотно расшифрована фотография».
Я: «Так от подобной ошибки суть сохранности остеоцита тираннозавра в течение «65 млн. лет» не изменяется».
П.: «Вы представляете себе, что такое «миллион лет»? Нет, вы не можете себе это представить, поскольку мы с вами живем в других временных измерениях. Что там может сохраниться за миллион лет?»
Я: «Так вы отрицаете длительные временные промежутки в истории Земли?»
П.: «Я ничего не отрицаю, просто говорю, что миллионы лет сохранности таких препаратов мне непонятны».
Я: «А вот для мягких тканей показано окаменение, когда они минералами пропитываются. Могут они долго сохраняться тогда?»
П.: «Наверное, могут, если пропитаются».
Я: «И что, тогда гибкие прозрачные сосуды, эритроциты и клетки костного мозга в минерализованном образце миллионы лет выдержат? Что их потом можно будет под микроскопом идентифицировать?»
П.: «Не знаю, непонятно, как такое может быть, невозможно представить. Про миллионы лет говорили уже...»
Я: «А в тех статьях различные гипотезы выдвигаются, которые как раз сохранность клеток и тканей в течение миллионов лет объясняют...».
П.: «Ну, вы сами должны понимать, что объяснить все можно...».
Я: «Тогда дайте мне какую-нибудь ссылку на время сохранения у трупа костного мозга. Наверное, у вас есть что-нибудь по судебной медицине в этом роде».
П.: «Таких ссылок нет и не ищите, поскольку данный вопрос в судебной медицине никому не интересен. Важно идентифицировать костный мозг у трупа на предмет предполагаемых патологий или токсических воздействий, а сколько он может храниться, никого так прямо не волнует».
Должен заметить, что мой опрос вместе с показом фото длился быстро — за один раз, и наш патологоанатом не имел достаточного времени для размышлений. Наиболее же важным результатом диалога является, во-первых, отсутствие сомнений у специалиста в достоверности изображенного на тех фото, и, во-вторых, в его исходной реакции на «миллионы лет» сохранности костного мозга, клеток и сосудов, которая, вероятно, и отражает реальную ситуацию.
6.3. Гипотезы о сохранности костного мозга «миллионы лет»: кость как
герметичный контейнер и самородная сера как стабилизатор
Конечно, костный мозг в окаменелых образцах может сохраняться много дольше, чем «десять дней в холодильнике». Просто в патологоанатомических случаях трупы, как правило, не попадают в столь благоприятные условия (почти без доступа воздуха, под наносами и т.п.). Возьмем, к примеру, исследование фоссилизованного костного мозга в останках людей эллинистической эпохи, обнаруженных в районе Персидского залива. Были даже идентифицированы эритроциты необычной формы — серповидной (серповидноклеточная анемия) [180]. Ясно, однако, что эллинистическая эпоха — это не миллионы лет назад; кроме того, структуры костного мозга людей сохранились, вероятно, из-за сухого климата пустыни.
Но лягушки и саламандры в пустынях не живут; их останки с костным мозгом были найдены в Испании в отложениях древнего субтропического озера. По мнению руководителя исследований М. Макнамары, сохранение костного мозга с органическими остатками было обусловлено особенностями строения костей у амфибий. Поры в их костях столь малы, что пропускают воду, но не бактерий разложения. Вот поэтому-то клетки костного мозга и сохранились до сих пор [168]. Дескать, «кости действовали как защитная раковина» [174].
Странно думать, что кости даже амфибий в течение миллионов лет были подобны герметичному контейнеру. Да и как будто дело только в микробах, а не в химических факторах, радиации, температуре и пр. Подобная гипотеза, озвученная М. Макнамарой, может только удивить.
Но это еще не все: на зарубежном атеистическом сайте опубликована полемика по данному вопросу [181]. Автор соответствующей статьи в «New Scientist» пишет, что он провел определенное исследование, которое показало, что окаменелости были обнаружены в старых серных шахтах Испании, и что «сера просачивалась из воды в кость, изменяя состав органического материала. Сера замедляет распад органики. К тому же она делает ее более жесткой» [181].
По этому поводу нами также было проведено исследование — сетевое — на предмет того, в каком химическом виде могут быть месторождения серы (sulfur mine), в частности, в том месте (восточная Испания; Арагон; Libros, Teruel [168, 173]). Оказалось, что твердые месторождения серы бывают как в самородном виде, так и в виде солей — сульфидов, сульфатов и сульфосолей [182]. В тех месторождениях Испании, для которых мы смогли найти сведения (северная Испания), сера представлена в самородном виде [183]. Скорее всего, и в Арагоне сера — в самородном виде, что в частности следует из контекста публикаций [168, 181].
Мы должны сразу аннулировать гипотезу о том, что сульфиды, сульфаты и пр. (соли отнюдь не слабых кислот), растворившись в воде, могут как-то стабилизировать белки, липиды и липопротеины до такой степени, что в результате сохранится морфология состоящих из них клеток. Белки могли только денатурировать, и клеточные структуры разрушились бы.
Остается самородная сера, которая плавится до жидкого состояния при температурах более 110°C (в таких случаях проблема сохранения костного мозга, понятно, становится пустой). В кристаллическом виде сера нерастворима в воде, а растворима в полярных растворителях типа сероуглерода [184], но те в недрах Земли явно отсутствуют. Если же в том месте и произошел когда-то выброс чего-то вроде сероуглерода в связи с извержением вулкана, то, опять же, в подобном случае вопрос сохранности там клеток костного мозга вновь приобретает умозрительный характер.
Автор публикации в «New Scientist» [181], вероятно, слышал когда-то в школе, что в состав белков входит много серы, а S-S-мостики в молекулах белка придают им структуру и жесткость. Все это так, но чтобы сера вошла в состав белка животных, необходим, во-первых, источник органической серы (аминокислота цистеин, трипептид глутатион и пр.), и, во-вторых, специальные ферменты и белки, которые способны данную серу утилизировать (в мертвых клетках и тканях они не работают). Но даже в таком случае устойчивость к распаду и перевариванию бактериями у белков вряд ли кардинально повысится. Так что механизм стабилизации органических структур за счет их пребывания возле самородной серы [181] является просто измышлением.
И мы пока не имеем никаких реальных гипотез, способных объяснить сохранение костного мозга в течение «10 млн. лет». Которых (миллионов лет), впрочем, и не было вовсе.
7. КЛЕТКИ, ОСТАТКИ КОСТНОГО МОЗГА, МЫШЦ, БЕЛКОВ И ПРОЧЕГО НА КАМЕННЫХ
ОРУДИЯХ ВОЗРАСТОМ ДО «ДВУХ МИЛЛИОНОВ ЛЕТ»
Это относительно мало известный в области молекулярно-клеточной палеонтологии пласт исследований с удивительными результатами, которые долгое время подвергались критике, но так, похоже, и не опровергнуты. А мало известен он, поскольку входит, скорее, в область молекулярной археологии, и основная масса соответствующих публикаций имела место в журналах археологического профиля — «Antiquity», «Journal of Field Archaeology», «World Archaeology» и пр.
Смысл заключается в том, что на доисторических каменных орудиях, на обрывках древних шкур и пр. путем микроскопирования и иммунохимии ищут остатки доисторических биомолекул и даже биоструктур, включая клетки. Первому подобная мысль пришла в начале 1980-х гг. доктору Томасу Лою (Thomas Harold Loy), молекулярному археологу из Университета в Квинсленде, Австралия [185–187]. Это очень известная фигура в данной области, и его публикации 1980-х гг. цитировались доктором М. Швейцер по крайней мере в статье 1997 г. [55] и в обзоре по молекулярной палеонтологии [8]. Первая работа доктора Т. Лоя по идентификации остатков крови (гемоглобина) на доисторических орудиях возрастом в 1–6 тыс. лет увидела свет в 1983 г. в журнале «Science» [188], хотя далее пошли статьи уже в археологических журналах. Труды посвящены идентификации фрагментов крови и белков [189–193], причем в публикации об этих исследованиях за 1995 г. [194] упоминалось, что доктор Т. Лой был «одним из немногих, кто видел на каменном орудии остатки эритроцитов». Работы Т. Лоя с соавторами с успешными результатами продолжались с 1980-х [188–190] по 1990-е гг. [191–194] и закончились только недавно [195–197], в связи с его смертью (2005 г. [187]).
Отдельные аналогичные данные были получены и другими известными авторами: доктором Дж.М. Ловенштейном (J.M. Lowenstein) из университета в Сан-Франциско [194, 198] и доктором Норин Туросс (Noreen Tuross) из Смитсоновского института в Вашингтоне, которая считает, что идентифицировала остатки гемоглобина мастодонта на каменном орудии из Чили возрастом в 11.000 лет [194].
С самого начала корректность результатов нахождения остатков крови и, иммунохимически, соответствующих белков (альбумина [191, 195], иммуноглобулина [191, 194] и гемоглобина [188, 194]) на орудиях возрастом до 90.000 [191] и 100.000 [189] лет подвергалась большому сомнению. Как и мы, как и П. Палмквист в подразделе 4.4., критики отсылали к судебно-медицинским исследованиям, демонстрировавшим, что остатки крови не определяются в образцах уже спустя месяцы — десятки месяцев [148, 194]. Проводились даже специальные работы, когда древние каменные орудия с пятнами свежей крови захоранивали в почве. Оказалось, что в сухой почве кровь на орудиях определяется иммунохимическими методами не более чем через 10 месяцев, а во влажной — не более чем через месяц. Были расхождения и при исследовании одних и тех же образцов различными аналитическими группами вслепую и т.п. [194]. Словом, многих биохимиков убедить так и не удалось как в 1980-х и 1990-х гг. [194], так и в 2000-х гг. [195]. Но строгое опровержение, что получены фальшивые результаты и артефакты, нам так и не встретилось. Зато на университетском сайте Квинсленда [185, 186] и в австралийских журналах (например, [197]) имеются подробные данные про доктора Томаса Лоя, которые убеждают нас, что это не жулик. И уже было сказано, что доктор М. Швейцер неоднократно цитировала его работы [8, 55].
Вкратце основные вехи научной биографии Т. Лоя следующие (2004 г.): молекулярный археолог, доктор наук (Ph.D.), сотрудник университета в Квинсленде, Австралия, член Археологических обществ Австралии и США, председатель организационного комитета 7-й Международной конференции «Древняя ДНК и биомолекулы» («Ancient DNA and Associated Biomolecules»), состоявшейся в 2004 г. в Брисбене, Австралия [185–187, 197].
Отдав должное послужному списку ведущего молекулярного археолога покойного доктора Томаса Лоя, и убедившись, что жульничество ему вряд ли было свойственно, мы теперь смело переходим к его главной находке. Ее мы рассматриваем по публикациям на сайте университета в Квинсленде 1998 г. [186], в австралийском журнале 2004 г. [197] и в «Science Online» 2004 г. [195]. Соответствующий доклад доктора Т. Лоя был представлен на 7-й Международной конференции по древним ДНК и биомолекулам в Брисбене, Австралия (2004 г.) [196].
В конце 1990-х гг. доктор Т. Лой с соавторами проводил раскопки на месте стоянки древних гоминид близ Йоханнесбурга, Южная Африка («2 млн. лет»). В материале [195] имеется фото «Олдувайского» кварцевого ручного рубила из раскопа, причем оцененный возраст его составляет, понятно, «два миллиона лет» (рисунок 8). Были обнаружены и другие кварцевые орудия (порядка 2000 экземпляров), а на многих из них — двухмиллионолетние кровь и жир. «Я исследовал орудия, и всюду имелась кровь», — свидетельствовал доктор Т. Лой. Он обнаружил интактные и фрагментированные эритроциты и лейкоциты, жировые клетки костного мозга, коллаген связок и сухожилий, мышечные ткани и остатки шерсти. Были на орудиях также фрагменты дерева, кератин от обломков рогов и семена с крахмалом, «принадлежащие растению, все еще растущему в этой области» [186, 195, 197]. Всему — «два миллиона лет». Две из трех цитированных нами публикаций — 2004 г. [195, 197], когда, вероятно, южноафриканские результаты стали более известны благодаря 7-й конференции по древним биомолекулам [196].
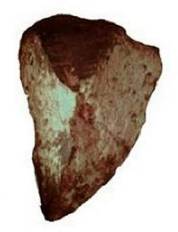
Рисунок 8. Кварцевое ручное рубило из Южной Африки — из
орудий возрастом в «два миллиона лет», на которых доктор Томас Лой из Австралии
обнаружил в 1998 г. остатки эритроцитов, лейкоцитов, клеток костного
мозга, фрагменты мышц, белков, семена и прочее [195].
Необходимо отметить, что доктор Т. Лой выработал некую собственную методику исследования, отличавшую его от других авторов. Если иммунохимики обычно просто промывали каменные образцы («экстрагировали камень») и искали все в промывных жидкостях (зачастую безрезультатно [194]), то Т. Лой, как он отмечал в [195], сначала пытался найти под микроскопом те места на орудиях, где могли быть основные остатки; иной раз он даже делал точные копии образцов и обрабатывал ими различные биоматериалы и ткани, чтобы узнать, где локализуются и как ведут себя органические остатки на рабочей поверхности орудия. И только потом приступал к поискам древней крови и прочих биоструктур в местах, так сказать, их «наибольшей ожидаемой концентрации» [195].
Согласно мнению доктора Т. Лоя, белки и остатки крови настолько хорошо закрепляются на каменной поверхности, что способны выдержать «миллионы лет», тем более в сухой почве [186, 195]. Ему, однако, указывали даже в 2004 г., что фрагменты белков не могут сохраняться в климате Южной Африки более чем 100.000 лет [195] (эту магическую величину что для ископаемых белков, что для ископаемых ДНК, что для Африки, что для Северной Америки и пр., мы подробнее рассмотрим ниже).
Следует отметить, что нахождение остатков крови, костного мозга (вплоть до клеток), фрагментов мышц, белков и прочего на каменных орудиях довольно серьезно подрывает самые главные из предложенных молекулярными палеонтологами механизмов сохранении биомолекул в течение геологических промежутков времени (см. подраздел 4.3). Ведь здесь нет никаких «стабильных комплексов с апатитом минерализованных костей». Здесь, в сущности, нет ничего, кроме каменной поверхности (у кварца вряд ли пористой) и биомолекул. И даже почва, вероятно, присутствует не всегда. Например, некоторые орудия с эритроцитами, альбумином, коллагеном и пр. возрастом в «90 тыс. лет» найдены в пещере Табун в Израиле, хотя, правда, и сказано, что там они тоже были «раскопаны» (excavated) [191].
Итак, ведущий молекулярный археолог, австралийский доктор Томас Лой с соавторами находил биоструктуры на каменных орудиях с 1983 г. [188] по конец 1990-х гг. [186, 193]. И явно находил бы их далее, все более погружаясь в глубь «миллионолетий»[19], если бы не его безвременная смерть. Некоторые другие молекулярные палеонтологи и археологи тоже обнаруживали биоструктуры на орудиях, хотя и в меньших масштабах [194, 198]. Нигде не видно сведений насчет ошибок и фальшивок, более того, все цитированные авторы — весьма уважаемые и известные исследователи. Даже если иммунохимические методы и привели к артефактам применительно к белкам, то как быть с клетками под микроскопом? И как мы должны все это интерпретировать, чтобы не впасть в полный маразм, поверив в эритроциты и клетки костного мозга, сохранившиеся на поверхности кварца (пусть даже в трещинах) в течение «двух миллионов лет»? Только так, что всему этому не «два миллиона» [186, 195–197], и не «100.000» [189], и даже не «90.000» [191] лет, а неизмеримо меньше.
8. ОРГАНИЧЕСКИЙ ЗАПАХ ОТ
ОКАМЕНЕЛЫХ КОСТЕЙ С АДСКОГО РУЧЬЯ
Факты, представленные в предыдущем разделе, уже не укладываются в голове даже в свете прочих находок. Но и они посрамлены: данные, приведенные здесь, выглядят вовсе анекдотом. Автор настоящего обзора не берет на себя никакой ответственности, а просто указывает ссылку: относительно недавняя обширная статья в «Discover» от апреля 2006 г. с интервью и рассказами доктора М. Швейцер [163].
«Когда М. Швейцер исследовала скелет тираннозавра, найденный в Адском Ручье[20], она заметила, что окаменелость источает отчетливо органический запах (organic odor). «Она пахла точно так же, как один из трупов, которые были у нас в лаборатории. Тот человек, прежде чем умер, подвергался химиотерапии» — говорит Швейцер. Учитывая обычное понятие, что подобные окаменелости составлены полностью из минералов, Швейцер обеспокоено обратилась к профессору Дж. Хорнеру. «Но он сказал в ответ только, что все кости с Адского Ручья пахнут (smell)» — вспоминает Швейцер. Старинные палеонтологи трупный запах (the smell of death) даже не регистрировали. Для Швейцер же этот факт подразумевал, что следы жизни могли все еще цепляться за те кости» [163].
Отметим тут, что к химиотерапии запах, скорее всего, отношения не имеет. Труп, что после химио-, что после радиотерапии, что без них, должен в целом пахнуть одинаково. Наверное, доктор М. Швейцер просто задумчиво вспоминала неприятные подробности, связанные с тем трупом, а корреспондент записывал все подряд.
Но, все-таки, не до конца ясно, что за запах источали кости тираннозавра. Это надо выяснить, прежде чем гнаться за сенсацией. Ведь из представленной цитаты следует, что доктор М. Швейцер упоминает какой-то «органический запах», в то время как про «трупный запах» написал сам автор статьи в «Discover», вместе со своими комментариями. Нельзя полностью исключить, что кости пахли не тлением, а какой-то «химией» из отложений.
9. РАДИАЦИОННЫЙ АСПЕКТ МОЛЕКУЛЯРНО-КЛЕТОЧНОЙ ПАЛЕОНТОЛОГИИ
Частично мы попытались рассмотреть данную проблему в нашей последней публикации [3]. Правда, мысли о радиационном аспекте молекулярно-клеточной палеонтологии не приходили в голову автору настоящего обзора вплоть до 2006 г., пока в статье в «Discover» [163] не попались рассуждения доктора Джеффри Бада (Jeffrey L. Bada), геохимика из Института океанографии в Сан-Диего. Дж. Бада сказал по поводу находок сосудов и клеток в костях динозавров [32, 113, 135, 163] (см. выше раздел 5), что мягкая ткань не может переживать миллионы лет, и что все образцы доктора М. Швейцер с соавторами — это просто результат современных загрязнений. Доктор Дж. Бада утверждал, что радиация окружающей среды должна была полностью разрушить ткани динозавров. И прибавил к этому, что «кости поглощают уран и торий как сумасшедшие, и доза от внутреннего облучения уничтожит все биомолекулы» [163].
Мы тогда сразу же рассчитали дозы внешнего облучения ископаемых биоструктур, накопленные ими за приписанные тем или иным костям «миллионы — десятки миллионов лет» за счет радиационного фона нашей планеты (преимущественно от радона в глубинах земли и от прочей радиоактивности пород; за вычетом из фона космической составляющей) [3]. Получились дозы вплоть за десятков миллионов рад (десятков миллионов «рентген»)[21], что тогда показалось нам слишком много даже для коротких фрагментов белков в сухом виде, не говоря уже о сосудах, клетках и мягких тканях. И мы, как и доктор Дж. Бада из Калифорнии, сделали тогда вывод, что «радиация за миллионы лет должна стереть все биоструктуры» [3].
К сожалению, мы с доктором Дж. Бада несколько поспешили в своих выводах [3, 163][22].
Находясь под влиянием слишком обобщающих представлений[23], мы в начале 2007 г. провели весьма обширное специальное исследование радиационного аспекта в молекулярно-клеточной палеонтологии, которое в полном виде на март 2007 г. еще не было опубликовано. Помимо расчета накопленной дозы внешнего облучения за счет радиационного фона Земли, мы оценили и дозу внутреннего облучения биоструктур в ископаемых костях за счет поглощенных трансурановых элементов — урана, тория, а также продуктов их распада радона и торона. Из соответствующих публикаций ([204–224] и др.) обнаружилось, что ископаемые кости действительно, как отмечал доктор Дж. Бада [163], поглощают трансурановые элементы «как сумасшедшие» (встретились упоминания, что из некоторых скоплений костей динозавров даже добывали уран, в том числе на заре выполнения ядерной программы США).
Не вдаваясь в подробности, мы скажем здесь, что, взяв за основу концентрацию урана и тория (а также соответствующий уровень радиоактивности радона и торона) в тех ископаемых костях (динозавров из Монголии [225, 226]), где это было определено, мы рассчитали, какую дозу радиации за счет радона и торона могли накопить сосуды и клетки в костях динозавров доктора М. Швейцер [32, 113, 135, 163], костный мозг миоценовых лягушек М. Макнамары [173–175] и фрагменты различных белков возрастами вплоть до «десятков — сотни с лишним миллионов лет» (см. выше раздел 4). Эти дозы кажутся очень большими[24]. Однако дальнейшее наше исследование в областях фундаментальной радиационной биофизики и радиобиологии больших доз показало, что малые фрагменты белков вполне способны выдержать дозы не только в десятки, но даже в сотни миллионов рад («рентген»). Обнаружился значительный пласт публикаций 1960–1990-х гг., посвященных определению молекулярной массы белков и полипептидных структур в том числе в составе мембран методом радиационной инактивации (для сухих препаратов). Оказалось, что выведены даже эмпирические формулы зависимости степени инактивации/деструкции (применительно к ферментам/белкам) от дозы радиации (γ-излучение, электроны) и температуры (см., например, [231–233]). Странно, что геохимик и давний специалист по астробиологии доктор Дж. Бада из США настолько мало знал об этих данных, что позволил себе в свое время усомниться в сохранности не только ископаемых клеток и тканей, но даже остеокальцина динозавров. Доктор Дж. Бада и тогда утверждал, что радиация за десятки миллионов лет должна полностью «стереть» все остатки даже столь небольшого и стойкого белка, как остеокальцин [119].
Наши расчеты по упомянутым выше формулам показали, однако, что фрагменты белков размером в 2000–3000 Да («у.е.») при температурах 0–20°C вполне могут выдержать без полного распада 600–900 млн. рад (мегарад; млн. «рентген»). (Мы проверяли эти свои расчеты неоднократно.) Но почти такой же уровень доз должен был быть накоплен за «десятки — сотню с небольшим миллионов лет» за счет внешнего облучения от среднего радиационного фона Земли плюс внутреннего облучения при умеренном (конечно) поглощении костями трансурановых элементов.
Поэтому что касается белков, вернее, их реагирующих с антителами (либо иммуногенных) относительно малых фрагментов, то мы вынуждены здесь сделать вывод, что применительно к молекулярно-клеточной палеонтологии радиационный аспект вряд ли дает креационные аргументы. Другое дело — ДНК[25] и морфологически неизменные остатки клеток, мягких тканей и сосудов в костях динозавров. В этом случае оцененные накопленные за «десятки миллионов лет» дозы радиации (особенно от внутреннего облучения за счет поглощенных урана и тория) кажутся слишком большими, хотя никаких точных формул и расчетов (как для белков и их фрагментов) в данном случае привести пока нельзя.
Мы надеемся полностью опубликовать наше исследование радиационного аспекта молекулярно-клеточной палеонтологии (очень обширно, сложно и специально) в отдельном труде. Здесь же еще раз повторим, что, по нашему мнению, радиационный фактор для фрагментов ископаемых макромолекул (не ДНК) не является однозначным креационным свидетельством. Креационными свидетельствами являются здесь, вероятно, упоминаемые нами ранее физико-химический и бактериальный факторы [1–3]. Фрагменты полипептидов, не говоря уже о клетках и мягких тканях (точнее — «морфологически полностью идентичных им структурах»), не способны выдержать в условиях Земли (не в вакууме, не при абсолютном нуле градусов и пр.) «десятки — сотни миллионов лет».
10. ЗАКЛЮЧЕНИЕ[26]
Лет сто пятьдесят, если не более, палеонтологи и геологи считали, что ископаемые останки и остатки возрастами в «сотни тысяч — сотни миллионов лет» способны сохраняться только в виде «камня» (окаменелостей), хитина, панцирей, углеродистых отпечатков, угля, нефти и газа. Никому и в голову не приходило, что в костях динозавров можно найти фрагменты белков, клетки и ткани, причем сохранившиеся, скорее всего, на органическом уровне. И это несмотря на то, что палеонтологи прошлого часто похвально не сомневались в том, что возраст Земли соответствует Библии и насчитывает менее десятка тысяч лет.
Но представить себе, что фрагменты органических молекул способны выдержать «палеонтологические периоды», был не способен никто в течение долгого времени. Это не приходило в голову даже таким профессионалам научной фантастики, каким был профессор палеонтологии Иван Антонович Ефремов, мастер ископаемых находок. Мы помним его фантастические произведения на палеонтологические темы: «Звездные корабли», «Тень минувшего» и, наверное, некоторые другие. Но нигде профессор И.А. Ефремов и мысли не держал, что кто-то в костях динозавров найдет иммуногенные фрагменты белков самих динозавров. Не говоря уже о систематическом обнаружении сосудов и клеток, и костного мозга, возрастом в «10 млн. лет», и эритроцитов, лейкоцитов и жировых клеток на ручном рубиле в «2 млн. лет». Если бы про все такое кто-то сказал бы юному Ефремову в свое время, то, думаю, он и палеонтологом-то не захотел бы стать, поскольку это получается нечто вроде археолога-ветеринара. Что черепки от горшков, что кости от динозавров. Никаких, понимаете ли, «седых» миллионов лет, просто тысячелетия. Даже нам становится скучно, что уж говорить о палеонтологах и геологах...
Тем не менее, никто почему-то долго не пробовал, скажем, взять кость лет эдак «миллионов в семьдесят», получить из нее экстракт и хотя бы определить там белок. Каждый думал, что даже если белок и найдется, то только как результат современных загрязнений разной бактериальной и грибковой дрянью. Так, в своем первом обзоре по молекулярной палеонтологии за 2005 г. автор настоящего труда сравнивал подобные затеи с попытками выделить белок «из камней пирамид египетских или из булыжной мостовой» [1].
Но вот исследователь в области атомного оружия, физик Филипп Абельсон (P.H. Abelson) опубликовал в 1954–1956 гг. в американской периодике результаты своих изысканий в глубинах останков девонской рыбы (девонский период закончился «365 млн. лет» назад). Там обнаружились аминокислоты, которые посчитали принадлежащими той самой древней рыбе.
И вот, пожалуйста, положен исток кажущейся столь странной дисциплине, как молекулярная палеонтология. Если честно, то поначалу молекулярная палеонтология выглядит даже более странно, чем астробиология — поиск остатков органики и жизни в космосе и на других планетах. Но это так, только если принимать за правду построения о длительных геологических периодах времени в истории Земли.
Прошло порядка десяти лет, и в Польше, в университете Кракова, доктор Р. Павлички (R. Pawlicki) с соавторами увидел под микроскопом в кости динозавра возрастом в «80 млн. лет» клетки, фибриллы коллагена и сосуды. Статья была издана в 1966 г. в таком известном мiровом журнале, как «Nature» [19]. Затем в 1970-х — 1990-х гг. доктор Р. Павлички (практически без соавторов) опубликовал уже в журналах более узкого профиля и значимости 13 статей по нахождению остатков биоструктур в костях динозавров. Там были и эритроциты с железом, и остеоциты, и зарегистрированные иммуногистохимическими методами липиды, ДНК и углеводы. Именно доктор Р. Павлички из Кракова, по нашему мнению, является предтечей молекулярной палеонтологии, а не доктор Ф. Абельсон с теми аминокислотами возрастом не менее «365 млн. лет».
В 1970-х гг., помимо польских исследований, вышло еще несколько работ, среди которых наиболее яркой кажется статья по обнаружению довольно больших фрагментов гликопротеинов (порядка 10.000 Да — углеродных единиц) в раковине ископаемого моллюска возрастом в «80 млн. лет» [34].
1980-е гг. — дальнейшее развитие: в ископаемых костях (до «2 млн. лет» и «8 млн. лет») с помощью антител идентифицировали остатки альбумина и коллагена [37, 38]. А затем — и белка остеокальцина в «13 и 30 млн. лет» [39]. Начали публиковаться результаты исследований из областей молекулярной археологии и палеоантропологии, где детектировались остатки белков в костях исторических и доисторических людей. В 1984 г. с помощью полимеразной цепной реакции (PCR) впервые выделили ДНК вымершего животного — квагги (выбита в конце XIX в.). Этим было положено начало дисциплине палеогенетике, которая развивалась и развивается намного более бурно, чем молекулярно-клеточная палеонтология. Библиография по палеогенетике ныне насчитывает тысячи источников. В своем обзоре мы вопросы палеогенетики практически не рассматривали: это другая дисциплина, хотя накопленные в ее рамках данные совсем уж вопиюще свидетельствуют против «миллионолетних» возрастов ископаемых останков. Порядка двух десятков таких исследований 1990-х гг. ныне, в 2005 г., попросту признаны «неправильными», поскольку реанимированные фрагменты ДНК или бактерии насчитывают уж слишком много — «до сотен миллионов лет» [48].
Вероятно, в 1980-х гг. все уже начали помаленьку привыкать к тому, что в костях возрастами в «миллионы лет» сохраняются фрагменты белков, да не абы какие, а такие, что позволяют проводить их видоспецифичную идентификацию с антителами, когда определяется не просто полипептид как таковой, а конкретно альбумин, остеокальцин и пр. Привыкать все начали, но широкую известность молекулярная палеонтология в 1980-х гг., похоже, все еще не получила.
Настали 1990-е гг., которые принесли распространение новых методов (HPLC) и новые находки. 1992 г. — определение с антителами остатков остеокальцина в костях четырех динозавров возрастом от «73 до 150 млн. лет» [118–120]. Это уже кажется крайне странным, поскольку, чтобы антигенная детерминанта белка («эпитоп») реагировала с антителами, она должна состоять из не менее чем 6–8-ми аминокислот. Остеокальцин — малый белок, и подобная цепочка составляет порядка 12–16% от всей его молекулы, т.е., за «десятки — сотни миллионов лет» произошло уменьшение белка всего в 6–8 раз. Получается, что «период полураспада» остеокальцина как молекулы составляет от «24-х до 50-ти млн. лет». Берем молекулу белка остеокальцина, кладем и уходим. Возвращаемся через 24–50 млн. лет, и имеем нетронутую половину от исходной полипептидной цепи. Такая, понимаете ли, «научная реальность».
В 1990-е гг. нашли также коллаген и кератин в костях возрастами в «десятки миллионов лет», но самой известной оказалась работа по выделению иммуногенных фрагментов гемоглобина из кости тираннозавра возрастом в «65 млн. лет» — экстракт из кости индуцировал у крыс образование антител, отчетливо реагировавших с современными гемоглобинами [55]. С ископаемого гемоглобина начала свою работу группа ведущих молекулярных палеонтологов из университетов Монтаны и Северной Каролины во главе с профессором Джеком Хорнером (Jack Horner) и под непосредственным руководством ассистента профессора, доктора Мэри Швейцер (M.H. Schweitzer). Далее большинство основных находок в области молекулярно-клеточной палеонтологии связывают с последним именем.
В 1990-х гг. доктор М. Швейцер с соавторами обнаружили, помимо остатков гемоглобина, в кости того тираннозавра отчетливо видимые под микроскопом эритроциты с ядрами, но осмелились опубликовать эти данные только в научно-популярном журнале «Earth» за 1997 г. [44]. Даже и за эту жалкую публикацию они были подвергнуты жесткой критике, поскольку никто не верил, что клетки могут сохраняться в костях динозавров, а про аналогичную работу польского доктора Р. Павлички за 1966 г. [19], вероятно, забыли.
Креационные авторы (как, например, доктор К. Виланд) живо откликнулись на гемоглобин и эритроциты тираннозавра в своих публикациях, за что были подвергнуты критике эволюционистами, которые указывали, что то и не эритроциты вовсе, а «производные клеток крови, модифицированные тысячелетиями геологических процессов», и что там был не гемоглобин, а какие-то его жалкие огрызки, которых только-только хватило, чтобы индуцировать антитела (?), и что нечего тенденциозно преувеличивать [166].
Настал XXI век, и произошел кардинальный перелом. Антикреационные авторы, насколько мы можем судить, тон своих выступлений сильно сбавили. Помимо прочего, к 2003 г. были опубликованы английские результаты исследования кости игуанодона возрастом в «120 млн. лет», где каким-то чудесным образом сохранились не только фрагменты остеокальцина, но и полипептиды большой молекулярной массы — до 66.000 Да [121, 123]. Главный же перелом настал в 2005 г., когда в «Science» доктором М. Швейцер с соавторами были обнародованы результаты опытов над костями целых четырех динозавров — трех тираннозавров и гадрозавра. После растворения минеральной составляющей в образцах осталась только органическая компонента, которая была представлена мягкой тканью и отменно сохранившимися клетками. Под микроскопом были отчетливо видны гибкие, полые, прозрачные сосуды, частично сохранившие упругость при манипуляциях, а внутри сосудов — красные или коричневые эритроциты с ядрами. Кроме того, идентифицировали фибриллы костного матрикса и клетки кости — остеоциты с гибкими филоподиями-ножками [32, 114]. Приведенные в Интернете многочисленные цветные фотографии из оригинальных статей [32, 113] обошли, вероятно, половину научно-популярных сетевых источников.
И никто уже не критиковал доктора М. Швейцер за ее сенсационные клетки крови, и все свыклись с этим моментом. Более того, если сначала М. Швейцер, увидев под микроскопом гибкие сосуды и красные эритроциты динозавров, не поверила своим глазам, пока эксперимент не повторили 17 раз [171], то несколько позже она уже утверждала (2006 г.), что мягкие ткани сохраняются в очень многих костях динозавров, и их легко там найти, стоит только разломать [135].
Поиски мягких тканей и клеток в костях динозавров и других ископаемых животных действительно продолжались, и к 2007 г. были получены еще подобные данные, причем рекордом, видимо, следует считать такие структуры из кости гадрозавра возрастом в «80 млн. лет» [136]. Теперь М. Швейер любит показывать при выступлениях одновременно две микрофотографии образцов, приговаривая: «Одним из этих клеток 65 млн. лет, а другим — 9 месяцев. Можете вы сказать мне, какие из них какие?» [135].
В последней, сугубо научной статье доктора М. Швейцер с соавторами на начало 2007 г. реальность всех этих находок еще раз подтверждается [172].
Но не думайте, что одной только М. Швейцер все и исчерпывается. В августе 2006 г. в журнале Геологического общества Америки были опубликованы результаты обнаружения в останках миоценовых амфибий хорошо сохранившегося костного мозга, с клетками, сосудами, красными и желтыми кровяными ростками и т.п. Все было найдено в отложениях древнего озера в Испании возрастом в «10 млн. лет» [173]. И авторам работы [173], и нам, глядя на фото костного мозга, ясно, что в составе структур почти наверняка сохранились биомолекулы - в костях многих и многих лягушек и саламандр (и одного головастика) [173, 177] – все возрастом в «10 млн. лет»...
«Не дивитесь сему; ибо наступает время...» (Ин. 5:28).
И исследовательница костного мозга миоценовых амфибий, Мария Макнамара (M.E. McNamara), как и доктор М. Швейцер, ничуть не удивляется и считает, что сохранность этой ткани в ископаемых костях возрастом в «миллионы — десятки миллионов лет» дело как будто обычное, и дивиться тут нечему [173, 177, 178].
Поэтому мы смело поменяли прежний общепринятый термин «молекулярная палеонтология» [1, 2, 6–8] на «молекулярно-клеточная палеонтология», раз сохранность клеток возрастом в «миллионы лет» — дело как бы обычное. Мы впервые воспользовались этим термином начиная с января 2007 г. [3].
Теперь, после 2005–2006 гг., мы живем в новом времени, в новом измерении, где удивляться чему-либо не следует: подумаешь, гибкие, прозрачные сосуды и окрашенные клетки возрастом в «65–80 млн. лет». Видно, их столь невероятная стойкость, неведомая и оловянному солдатику — это закон природы и «медицинский факт»! Раз клетки и сосуды найдены в останках возрастом в «десятки миллионов лет», то, следовательно, они способны столько и выдержать. Такова логика, основанная на представлениях о длительных геологических эпохах, необходимых для макроэволюционных предрассудков.
Справедливости ради надо отметить, что «новое время», все-таки, мелькало еще в 1966 гг., когда польские авторы увидели в кости динозавра сосуды и клетки [19], а подкрепилось эритроцитами тираннозавра доктора М. Швейцер в середине 1990-х гг. [44]. В конце же 1990-х гг., после длительного экспериментального пути, клетки крови (эритроциты и лейкоциты), жировые клетки костного мозга, а также остатки мышц, шерсти и пр. были обнаружены на кварцевых орудиях из Южной Африки возрастом в «2 млн. лет» [186, 195–197]. Это поработал австралийский доктор Томас Лой (Thomas Harold Loy), который как раз и специализировался на поиске биоструктур на каменных орудиях начиная с 1983 г. [188]. И снова — нечего удивляться тому, что на кварце сохранились биоструктуры в течение «2 млн. лет». И снова — это закон природы и «медицинский факт».
А мы-то — и не знали, что живем в «новое время», узнали же об этом только в XXI веке, когда прозрачные сосуды и красные эритроциты динозавров полезли изо всех научно-популярных щелей, а не только из фильмов Стивена Спилберга и «BBC — живой мир».
Понятно, что крайне интересно, как же объясняет «наука» сохранность плохо сохраняемого в течение таких временных промежутков, когда и от хорошо сохраняемого-то мало что должно сохраняться. Опубликован ряд гипотез типа формирования специфических устойчивых биополимеров, образования комплексов с неорганическими соединениями почвы или быстрого «цементирования» биоструктур при захоронении в составе неких «внутренних кристаллов». Рассматриваются вопросы крайнего повышения устойчивости биомолекул в виде их комплексов с апатитом костей [7, 8, 125–134]. Но все такие гипотезы, несмотря на несомненные зерна истины, не способны объяснить столь длительные сроки «жизни» и устойчивость к микробным факторам. Теория же о сохранности костного мозга в костях амфибий, озвученная М. Макнамарой, согласно которой поры в этих костях столь малы, что не пропускают микробов и, поэтому, ткань сохранилась «10 млн. лет» [168, 174], понятно, никакой критики не выдерживает. Равным образом не выдерживает критики и мысль доктора М. Швейцер, что индуцированные ионами железа из распавшегося гемоглобина свободные радикалы способны стабилизировать белки путем формирования внутри- и межмолекулярных сшивок [135, 136]. От этого, дескать, и сохраняются гибкие прозрачные сосуды, красные эритроциты и остеоциты с филоподиями. Но гипотеза совершенно неверна: катализируемая ионами двухвалентного железа реакция Фентона, когда образуется крайне активный гидроксильный радикал, способна только разрушать биомолекулы вплоть до низкомолекулярных соединений, но не стабилизировать.
К тому же, если сравнить степень идентификации в ископаемых костях стабильных белков (кератина, коллагена и остеокальцина) с сохранностью таких лабильных белков, как гемоглобин, глико- и фосфопротеины, а также растворимые полипептиды, то никакого преимущества у первых нами обнаружено не было. Данные, сведенные здесь в две обширные таблицы, отчетливо указывают нам, что и те, и другие белки способны сохраняться вплоть до «десятков — сотни миллионов лет». Это странно.
Еще более странно то, что исследованные белки в археологических и палеоантропологических останках возрастом порядка тысяч — «десятка тысяч лет» определяются, как правило, ничуть не лучше, чем в костях динозавров и пр., имеющих возрасты в тысячи — десяток тысяч раз большие... Понятно, что условия хранения могут быть разными, но не настолько же, чтобы продлевать срок жизни белков в тысячи раз. Причем подобная несуразность выявлена даже для устойчивых белков — коллагена и остеокальцина. В останках «неандертальцев» из пещер Ирака и Хорватии найти коллаген не удалось, но он обнаруживался в костях, насчитывающих «десятки миллионов лет». В останках бизонов возрастом в 120 и 300 тыс. лет уровень остеокальцина убывает по экспоненциальной зависимости, достигая к 300 тыс. лет просто следовых, ничтожных концентраций в кости [1, 98]. Но это не помешало остеокальцину определяться в различных исследованиях и разными авторами в костях шести (пока) динозавров возрастом от «73 до 150 млн. лет» [118–120] (!), и определяться одними и теми же радиоиммунологическими и иммунохимическими методами.
Совсем недавно, когда представленный вам обзор был уже практически завершен, группа доктора М. Швейцер в одной из изученных ранее костей тираннозавров с мягкими тканями (образец MOR 1125) идентифицировала остатки коллагена. Причем не только идентифицировала, но даже секвенировала. А именно: для семи фрагментов белка была расшифрована аминокислотная последовательность (sequence — последовательность). И мы вновь увидели соответствующие солидные статьи в авторитетном мировом журнале «Science», причем вновь, как и в 2005 г. [11, 32], сразу две [233а, 233б].
Образцу MOR 1125 был приписан возраст в «68 млн. лет» [233а–233е]. В научно-популярном эволюционном мире данная находка вызвала ажиотаж, как будто не было предыдущих почти 40 лет молекулярно-клеточной палеонтологии. И мы видим в публикации BBC News, что ископаемый коллаген 2007 г. «намного старше органических молекул, экстрагированных до этого из окаменелостей», и что «обнаружение белка в костях динозавра является сюрпризом — органический материал, как считалось, не может выдержать столь долго». И т.д., и т.п. [233г, 233д]. Полностью забыто, что в кости игуанодона, которой приписан возраст аж в 120 млн. лет, еще семь лет назад английскими исследователями с помощью ионообменной хроматографии и электрофореза были идентифицированы полипептиды порядочного размера — до 66.000 Да [121, 123] (см. выше подраздел 4.2). Судя по всему, в своем интервью BBC [233г] ни доктор М. Швейцер, ни профессор Джек Хорнер не упомянули английские работы группы Embery G. 2000 и 2003 гг. [121, 123].
Правда, в области исследования древних полипептидов указанные авторы из США действительно имеют приоритет: до сих пор никому не удалось расшифровать даже фрагменты аминокислотной последовательности белков возрастов в «десятки миллионов лет». Эти последовательности доктор М. Швейцер с соавторами сравнили с последовательностями коллагенов современных животных (по специальной мировой базе данных). Оказалось, что последовательности коллагена T. rex и цыпленка были сходны на 58%, в то время как с коллагенами лягушки и тритона имелось сходство «только» на 51% [233а, 233б, 233е]. Отметим тут, что расшифрованы последовательности не всего белка тираннозавра, а только семи его отдельных фрагментов, т.е. вполне вероятно, что вкупе с отсутствующими фрагментами эта последовательность коллагена тираннозавра может быть ближе не к птицам, а, к примеру, к крокодилам. Строго говоря, пока нельзя исключить и наибольшего сходства даже с коллагеном некого сторонника эволюционной теории.
В [233е] отмечается к тому же, что, согласно первоисточникам [233а, 233б], хотя некоторые пептидные фрагменты белка тираннозавра и показали большее сходство последовательностей с коллагеном цыпленка, другие последовательности были более сходны с параметрами для лягушки и тритона (даже для рыбы и мыши).
Вот на основе какого неполного материала ныне публикуются достаточно громкие утверждения о том, что эти данные доктора М. Швейцер с соавторами еще раз доказывают на молекулярном уровне эволюционное родство между динозаврами и птицами. Приведем названия только некоторых статей: «Белок, связывающий тираннозавра и цыпленка» [233г], «Белок доказывает родство королевского тираннозавра с современной курицей» [233д], «Тираннозавр сродни цыпленку» [233ж] и другие.
И никому, за исключением западных креационистов [233е], не пришло в голову сказать, что отдельные фрагменты — это еще не весь белок, и что при сравнении одного уникального не полностью сохранившегося образца с табельными параметрами современных белков сходства на 58% (с белком цыпленка) и на 51% (с белками земноводных) нельзя рассматривать как разную степень отличия (т.е., те 58% и 51% на самом деле не являются точными, окончательными величинами, как в математике). Кроме того, у нас есть все тот же аргумент: сходство даже на молекулярном уровне различных животных никак не доказывает их происхождение от одного эволюционного предка (ибо у нас всегда есть пример с карбюраторными двигателями «Оки» и «Урала» — единый план творца).
Итак, в 2007 г. мы вновь узнали о нахождении значительных фрагментов коллагена с приписанным возрастом в «68 млн. лет». И это — вкупе с предыдущими данными по идентификации коллагена иммунохимическими методами в образцах, также насчитывающих «десятки миллионов лет» (см. выше табл. 2 в подразделе 4.2). Но, повторимся, данный белок нередко не удается обнаружить в образцах «моложе» на три порядка. И действительно, согласно термодинамическим расчетам самих молекулярных палеонтологов за 2002 г. (ранее неизвестная нам важная научная публикация [233з]), период полураспада даже стойкого коллагена при хранении в условиях 0°C составляет максимум 2,7 млн. лет. Если же белок выдерживать при 10°C, то этот показатель сокращается до 180 тыс. лет. А при 20°C — до 15 тыс. лет [8]. Следовательно, даже если тот тираннозавр MOR 1125 все время хранился бы на тающем льду (для штата Монтана невероятно), за 68 млн. лет он претерпел бы более 25-ти периодов полураспада, а именно — исходное количество коллагена разрушилось бы в 225 раз. Это — в 34 млн. раз, какие уж тут аминокислотные последовательности. (Отметим, что период полураспада еще более стойкого остеокальцина при указанных выше температурах составил бы, по расчетам [233з], 110 млн. лет, 7,5 млн. лет и 580 тыс. лет соответственно.)
Выходит так, что какие-то из молекулярно-клеточных палеонтологов ошибаются: или те, кто насчитал невероятное количество периодов полураспада для белков динозавров с оцененными возрастами в «десятки миллионов лет» [233з] (главная исследовательница ископаемых остеокальцинов [98, 152]), или те, кто приписал этим останкам указанные «миллионы лет». Выбирайте сами, какие палеонтологи более заслуживают вашего доверия.
Все эти «чехарды» сроков сохранности белков, постоянно и закономерно мало отличающиеся что для тысяч, что для «десятков миллионов лет», подкрепляют мысль, согласно которой с датировкой ископаемых останков имеется какая-то несуразность -не говоря уже о «систематическом» обнаружении морфологически неизменных сосудов, эритроцитов и остеоцитов в костях динозавров.
Помимо прочего, для ископаемых костей показано очень активное поглощение трансурановых элементов — радиоактивных урана и тория, которые со временем распадаются до радиоактивных радона и торона. Наше специальное исследование (в марте 2007 г. еще не было опубликовано) показало, что ископаемые кости динозавров, как правило, представляют собой нечто вроде малых «ядерных реакторов», так, что палеонтологи во всем мiре ищут эти кости в метре под землей с помощью счетчиков радиоактивности, а из некоторых скоплений костей динозавров добывали и добывают уран, в том числе делали это на ранних этапах выполнения ядерной программы США [204–224] (и др.).
Поэтому в области молекулярно-клеточной палеонтологии мы никак не можем сбрасывать со счетов и радиационный фактор, о чем свидетельствуют и другие исследователи, например — геохимик и астробиолог доктор Дж. Бада из США, который поэтому вовсе отрицает реальность каких-либо находок «палентологческих» ископаемых биоструктур, причем не только клеток и тканей [163], но даже малых фрагментов белков [119].
Мы оценили дозу внешнего (за счет радиационного фона Земли) и внутреннего (за счет умеренного накопления урана и тория костями) облучения сосудов, клеток, тканей и фрагментов белков, которые они должны были накопить за приписанные тем или иным костям «десятки — свыше сотни миллионов лет». Оказалось, что, если исходить из формулы дозовой зависимости при определении молекулярной массы сухих белков методом радиационной инактивации при 0–20°C [231–233], то не слишком большие полипептиды действительно могут выдержать подобную лучевую нагрузку. Однако для сохранившихся морфологически неизменными клеток и мягких тканей (точнее — для «полностью идентичных им морфологических структур») это кажется очень маловероятным.
Все эти несуразности с накопленными дозами радиации, с неизбежным воздействием физико-химического и почти наверняка (за «миллионы лет»!) бактериального факторов, с «чехардами» сроков одинакового сохранения для фрагментов одних и тех же белков, но по возрасту отличных на много порядков, кажутся весьма странными в рамках общерпринятых представлений о «многомиллионолетних» геологических эпохах. И только если мы примем библейское положение о том, что никаких «миллионов лет» просто не было, все встает на свои места.
Мы убеждены в этом и верим, что именно Книга Бытия, а вовсе не современные пособия по геологии и палеонтологии, в целом ближе к пониманию возраста Земли. Наша вера подкреплена соответствующими исследованиями. И нам непонятно, в особенности после всего изложенного в обзоре, как считающие себя христианами, причем православными, могут находиться на позициях теологического эволюционизма [234–240]. Мало того, что их «эволюционизм», в самом деле, это нечто вроде явной ереси [241–254], он еще и не верен с научных позиций. Глупость какая-то: чтобы угодить «науке» и выглядеть «научнее», такие христиане, теряя суть христианства — искупление Христом греха Адама [241, 242, 246], настоящей, корректной научности, оказывается, и не приобретают. Вполне понятно: утеряв Самую Большую Истину, неизбежно потеряешь и все меньшие.
«Смотрите, братия, чтобы кто не увлек вас философиею и пустым обольщением, по преданию человеческому, по стихиям мiра, а не по Христу...» (Кол. 2:8).
«...но осуетились в умствованиях своих, и омрачилось несмысленное их сердце; называя себя мудрыми, обезумели» (Рим. 1:21, 22).
«И еще: Господь знает умствования мудрецов, что они суетны» (1 Кор. 3:20).
Почему так — не совсем понятно, хотя есть богословское мнение, что у ученых психика очень уж следует обыденно-научным штампам [255], которые забивали в голову с детских лет [256]. Не знаю...
Как бы там ни было, некультурно это у них, у теологических эволюционистов, получается [234–240], хотя диакон Андрей Кураев и любит учить [236], а мы за ним повторять [3], что всегда следует разговаривать исключительно на «языке науки и культурологии».
Поэтому мы надеемся, что наш обзор, изложенный почти сугубо на «языке науки» (полагаем также, что и культурно), сможет поддержать колеблющихся, порадовать устойчивых, поразить несоглашающихся и, в конце концов, послужить одной из опор при формировании научной базы истинно православного мiровоззрения.
Аминь.
СПИСОК ЛИТЕРАТУРЫ
Примечание. Отметка «Есть сетевая версия» означает, что в
Интернете имеется в свободном доступе бесплатная электронная копия. Платные же
версии есть для большинства научных журналов за последние годы.
2. Лунный А.Н. Вслед за гемоглобином тираннозавра — мягкие ткани с эластичными сосудами и ядерными клетками из костей четырех динозавров. И вновь — фрагменты белков. В кн.: «Православное осмысление творения мiра». Выпуск 2. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», Москва, 2006. С. 179–202. (Есть сетевые версии.)
3. Лунный А.Н. Костный мозг, хранившийся «10 миллионов лет», еще кости динозавров с сосудами и эритроцитами, запах от останков возрастом «около 70 миллионов лет», мумии динозавров и прочее. Находки становятся обыденностью. В кн.: «Православное осмысление творения мiра». Выпуск 3. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», М.: Изд-во «Шестодневъ», 2007. С. 156–201. (Есть сетевые версии.)
4. Православный сайт «Слово отеческое». Сотворение мiра — данные современной науки (http:// www.slovotech.nm.ru/.)
5. Лунный А.Н. Веб-страница в библиотеке «Электронные полки Вадима Ершова» (публикации в архивах). 2006. (http://publ.lib.ru/ARCHIVES/L/LUNNYY_A._N/_Lunnyy_A._N..html.)
6. Розанов А.Ю. Современная палеонтология // Соросовский образовательный журнал. 1999. № 1. С. 47–55. (Есть сетевая версия.)
7. Стоник В.А. Молекулы
свидетельствуют о прошлом // Соросовский образовательный журнал. 2001.
Т. 7. № 3. С.
18–24. (Есть сетевая версия.)
8. Schweitzer M.H. The future of Molecular Paleontology //
Palaeontologia Electronica. 2003. V. 5. № 2. (http://palaeo-electronica.org.)
9. Schweitzer M.H., Watt J.A., Avci R. et al. Beta-keratin specific
immunological reactivity in feather-like structures of the cretaceous
alvarezsaurid, Shuvuuia deserti // J.
Exp. Zool. (Mol. Dev. Evol.). 1999. V. 285. № 2 . P. 146–157.
10. Schweitzer M.H., Marshall C.L. A molecular model for the evolution
of endothermy in the theropod-bird lineage // J. Exp. Zool. 2001. V. 291. № 4.
P. 317–338.
11. Schweitzer M.H., Wittmeyer J.L., Horner
J.R. Gender-specific reproductive tissue in ratites and Tyrannosaurus rex // Science. 2005. V. 308. № 5727. P. 1456–1460.
12. Currat M., Excoffier L. Modern humans did not admix with
Neanderthals during their range expansion into Europe // PLoS Biology. 2004. V.
2. № 12. P. 2264–2274. (Есть сетевая версия.)
13. Green R.E., Krause J., Ptak S.E. et al. Analysis of one million base
pairs of Neanderthal DNA // Nature. 2006. V. 444. № 16. P. 330–336. (Есть сетевая версия.)
14. Rogaev E.I., Moliaka Y.K., Malyarchuk B.A. et al. Complete
Mitochondrial Genome and Phylogeny of Pleistocene Mammoth Mammuthus primigenius // PLoS Biology. 2004. V. 4. № 3. P. 403–410.
(Есть сетевая версия.)
15. Beja-Pereira A., Caramelli D., Lalueza-Foxe C. et al. The origin of
European cattle: Evidence from modern and ancient DNA // Proc. Natl. Acad. Sci.
U.S.A. 2006. V. 103. № 2. P. 8113–8118. (Есть сетевая версия.)
16. Abelson P.H. Paleobiochemistry // Carnegie Inst. Washington Yearb. 1954. V. 53. P. 97−101; Scient. Amer. 1956. V. 195. P. 83–92. (См. также (http://ucnuclearfree.org/articles/2004/08/09_schudel_atom-bomb-scientist-dies.htm).
17. De Jong E.W., Westbroek P., Westbroek J.F., Bruning J.W. Preservation of antigenic properties of macromolecules over 70 Myr // Nature. 1974. V. 252. № 5478. P. 63–64.
18. Челомина Г.Н. Древняя ДНК // Генетика. 2006. V. 42. № 3. С. 293–309.
19. Pawlicki H., Korbel A., Kubiak H. Cells, collagen fibrils and
vessels in dinosaur bone // Nature. 1966. V. 211. № 49. P. 655–657.
20. Nowicki Z., Sarnecka-Keller M., Pawlicki R. et al. Morphological and
chemical analysis of the structure of the fossil dinosaur bone // Anat. Anz.
1972. V. 132. № 1. P. 10–23.
21.
Pawlicki R. Studies of the fossil dinosaur bone in the scanning electron
microscope // Z. Mikrosk. Anat. Forsch. 1975. V. 89. № 2. P. 393–398.
22.
Pawlicki R. Topochemical localization of lipids in dinosaur bone by means of
Sudan B black // Acta Histochem. 1977. V. 59. № 1. P. 40–46.
23.
Pawlicki R. Histochemical reactions for mucopolysaccharides in the dinosaur
bone. Studies on Epon- and methacrylate-embedded semithin sections as well as
on isolated osteocytes and ground sections of bone // Acta Histochem. 1977. V.
58. № 1. P. 75–78.
24.
Pawlicki R. Morphological differentiation of the fossil dinosaur bone cells.
Light, transmission electron-, and scanning electron-microscopic studies //
Acta Anat. (Basel). 1978. V. 100. № 4. P. 411–418.
25. Pawlicki R. Metabolic pathways of the fossil dinosaur bones. Part I.
Vascular communication system // Folia Histochem. Cytochem. (Krakow). 1983. V.
21. № 3–4. P. 253–261.
26. Pawlicki R. Metabolic pathways of the fossil dinosaur bones. Part
II. Vascular canal in the communication system // Folia Histochem. Cytobiol.
(Krakow). 1984. V. 22. № 1. P. 33–41.
27.
Pawlicki R. Metabolic pathways of the fossil dinosaur bones. Part III.
Intermediary and other osteocytes in the system of metabolic pathways of
dinosaur bone // Folia Histochem. Cytobiol. 1984. V. 22. № 2. P. 91–97.
28. Pawlicki R. Metabolic pathways
of the fossil dinosaur bones. Part IV. Modes of linkage between osteocytes and
a variety of nexuses of osteocytes processes // Folia Histochem. Cytochem. (Krakow).
1984. V. 22. № 2. P. 99–104.
29.
Pawlicki R. Metabolic pathways of the fossil dinosaur bones. Part V.
Morphological differentiation of osteocyte lacunae and bone canaliculi and
their significance in the system of extracellular communication // Folia Histochem.
Cytobiol. 1985.V. 23. № 3. P. 165–174.
30. Pawlicki R. Histochemical demonstration of DNA in osteocytes from
dinosaur bones // Folia Histochem. Cytobiol. (Krakow). 1995. V. 33. № 3. P.
183–186.
31. Pawlicki R., Nowogrodzka-Zagorska M. Blood vessels and red blood
cells preserved in dinosaur bones // Anat. Anz. 1998. V. 180. № 1. P. 73–77.
32.
Schweitzer M.H., Wittmeyer J.L., Horner J.R., Toporski J.K. Soft-Tissue Vessels
and Cellular Preservation in Tyrannosaurus
rex // Science. 2005. V. 307. № 5717. P. 1952–1955. (В
2005 г. была сетевая версия;
в 2006 г.
стала недоступна.)
33.
Armitage M. Scanning electron microscope study of mummified collagen fibers in
fossil Tyrannosaurus rex bone // CRSQ
(Creation Research Society Quarterly). 2001. V. 38. № 2. P. 61–66. (Есть сетевая версия.)
34.
Weiner S., Lowenstam H.A., Hood L. Characterization of 80-million-year-old
mollusk shell proteins // Proc. Natl. Acad. Sci. U.S.A. 1976. V. 73. № 8. P.
2541–2545. (Есть сетевая версия.)
35.
Westbroek P., Van der Meide P.H., van der Wey-Kloppers J.S. et al. Fossil
macromolecules from cephalopod shells: characterization, immunological response
and diagenesis // Paleobiology. 1979. V. 5. № 2. P. 151–167.
36.
Towe K.M., Urbanek, A. Collagen-like structures in Ordovician graptolite
periderm // Nature. 1972. V. 237. P. 443–445.
37.
Lowenstein J.M. Species-specific proteins in fossils // Naturwissenschaften.
1980. V. 67. № 7. P. 343–346.
38.
Lowenstein J.M. Immunological reactions from fossil material // Philos. Trans.
R. Soc. Lond. B. Biol. Sci. 1981. V. 292. № 1057. P. 143–149.
39.
Ulrich M.M., Perizonius W.R., Spoor C.F. et al. Extraction of osteocalcin from
fossil bones and teeth // Biochem. Biophys. Res. Commun. 1987. V. 149. № 2. P.
712–719.
40. Tuross N. Albumin preservation in the Taima-taima mastodon skeleton
// Appl. Geochem. 1989. V. 4. № 3. P. 255–259.
41. Ascenzi A., Brunori M., Citro G., Zito R. Immunological detection of
hemoglobin in bones of ancient Roman times and of Iron and Eneolithic Ages //
Proc. Natl. Acad. Sci. U.S.A. 1985. V. 82. № 21. P. 7170–7172. (Есть сетевая версия.)
42. Lowenstein J.M. Molecular approaches to the identification of
species // American Scientist. 1985. V. 73. P. 541–546.
43. Runnegar B. Molecular palaeontology // Palaeontology. 1986. V. 29. №
1. P. 1–24.
44. Schweitzer M.H., Staedter T. The real Jurassic Park // Earth. 1997.
V. 6. № 3. P. 55–57.
45. Loreille O., Kiesslich J. Ancient DNA references. 1999.
(http://www.comic.sbg.ac.at/staff/jan/ancient/aDNA%20library.html.) Ancient
Genome Encyclopedia (Ancient DNA DataBase). 2004.
(http://www.ddbj.nig.ac.jp/aDNA/index.html.)
46.
Discovers Soft Tissue In Dinosaur Bones. NC State Paleontologist //
ScienceDaily. March 25, 2005.
(http://www.sciencedaily.com/releases/2005/03/050325100541.htm.)
47.
Blin K. Paleogenetics. DNA analysis of ancient remains // Institute of
Anthropology and Human Genetics. University of Tubingen. Division of Molecular
Genetics. 1.08.2003.
(http://www.uni-tuebingen.de/uni/thm/molgen/paleogenetics.html.)
48.
Hebsgaard M.B., Phillips M.J., Willerslev E. Geologically ancient DNA: fact or
artefact? // Trends Microbiology. 2005. V. 13. № 5. P. 212–220. (Есть сетевая версия.)
49.
Woodward S.R., Weyand N.J., Bunnell M. DNA sequence from Cretaceous period bone
fragments // Science. 1994. V. 266. № 5188. P. 1229–1232.
50.
Li Y, An C-C, Zhu Y-X. DNA isolation and sequence analysis of dinosaur DNA from
Cretaceous dinosaur egg in Xixia Henan, China // Acta Sci. Nat. Univ.
Pekinensis. 1995. V. 31. P. 148–152.
51.
Hedges S.B., Schweitzer M.H. Detecting dinosaur DNA (есть сетевая версия);
Zischler H. et al. Detecting dinosaur DNA // Science. 1995. V. 268. № 5214. P.
1191–1193.
52.
Xu X., Wang X.L., Wu X.C. A dromaeosaurid dinosaur with a filamentous integument
from the Yixian Formation of China // Nature. 1999. V. 401. № 6750. P. 262–266.
53.
Qiang J., Currie P.J., Norell M.A., Shu-An J. Two feathered dinosaurs from
northeastern China // Nature. 1998. V. 393. № 6687. P. 753–761.
54.
Wuttke M. In: Messel-Ein Schaufenster in die Geschichte der Erde und des
Lebens, S. Schaal, W. Ziegler, Eds. (Verlag Waldemar Kramer, Frankfurt am Main,
Germany, 1988), pp. 265–274.
55.
Schweitzer M.H., Marshall M., Carron K. et al. Heme compounds in dinosaur
trabecular bone // Proc. Natl. Acad. Sci. U.S.A. 1997. V. 94. № 12. P.
6291–6296. (Есть сетевая версия.)
56.
Chiappe L.M., Coria R.A., Dingus L. et al. Sauropod dinosaur embryos from the
Late Cretaceous of Patagonia // Nature. 1998. V. 396. № 6708. P. 258–261.
57.
Dal Sasso C., Signore M. Exceptional soft-tissue preservation in a theropod
dinosaur from Italy // Nature. 1998. V. 392. № 6674. P. 383–387.
58.
Kellner A.W.A. Fossilized therapod soft-tissue // Nature. 1996. V. 379. № 6560.
P. 32.
59.
Schweitzer M.H., Horner J.R. Intravascular microstructures in trabecular bone
tissues of Tyrannosaurus rex // Ann.
Paleontol. 1999. V. 85. № 3. P. 179–192.
60. Briggs D.E.G., Wilby P.R., Perez-Moreno B.P. et al. The mineralization of dinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain // J. Geol. Soc. London. 1997. V. 154. P. 587–588.
61. Войткевич Г.В. Возникновение и развитие жизни на Земле. М.: Наука. 1988. — 144 с. (Есть сетевая версия.)
62. Баландин Р.К. Вернадский: жизнь, мысль, бессмертие. М: Знание, Серия «Творцы науки и техники», 1979. (http://lib.udm.ru/lib/filosof/wernadskij/balandin.txt_Contents.)
63.
Lines D. Fossilized Human Finger // The Creation Evidences Museum. 1995.
(http://home.texoma.net/~linesden/cem/finger/finger.htm.)
64. Guernsey’s (http://www.guernseys.com/Auctions/PastEvents/Dinosaur/Featured.html.)
65. Dino-Mummy Shows Some Skin
(http://www.sir-ray.com/Dino-Mummy%20Article.htm.)
66. Leonardo Project — What was Leonardo? Kodak Project
(http://www.kodak.com/eknec/PageQuerier.jhtml?pq-path=2608/2611/4141/9507/9611&pq-locale=en_US.)
67. Halls K.M. Dinosaur mummies: beyond bare-bone fossils. Illustr. by
R. Spears. Plain City, Ohio: Darby Creek Pub., 2003. — 48 p.
(http://catalog.sno-isle.org/cgi-bin/cw_cgi?fullRecord+19770+86+94335831+1+0);
(http://www.mummytombs.com/market/books/child/advpicturebooks/halls.dinosaur.htm.)
68. Fossil Facts
(http://www.mummydinosaur.com/MeetLeonardo/MeetFossil.htm;
http://dml.cmnh.org/2004Jul/msg00064.html.)
69. Иорданский Н.Н. Развитие жизни на Земле. М.: Просвещение. 1981. — 190 с. (Есть сетевая версия.)
70.
Dinosaur Mummy #1. Scipionyx samniticus
with preserved soft tissue.
(http://www.mummytombs.com/mummylocator/animal/dinosaur.htm.)
71. Scientists discover 66-million-year-old dinosaur with a heart.
Fossils in the News. Member of the American Federation of Mineral Societies.
Member of the Southeast Federation of Mineral Societies. 04.21.2000.
(http://www.gamineral.org/fossil-news.htm.)
72. Schweitzer M.H., Chiappe L., Garrido A.C. et al. Molecular
preservation in Late Cretaceous sauropod dinosaur eggshells // Proc. Biol. Sci.
2005. V. 272. № 1565. P. 775–784.
73. Murphy N.L., Trexler D., Thompson M. Exceptional soft-tissue
preservation in a mummified ornithopod dinosaur from the Campanian Lower Judith
River Formation. Society of Vertebrate Paleontology Meeting. 2002. Oct. 9–12.
Norman, Okla.
74. Dinosaur mummy sheds light on look, feel of prehistory.
«Scienceblog».
(http://www.scienceblog.com/cms/dinosaur_mummy_sheds_light_on_look_feel_of_prehistory.)
75. Mayell H. «Mummified» Dinosaur Discovered
In Montana // National Geographic News. October 11, 2002.
(http://news.nationalgeographic.com/news/2002/10/1010_021010_dinomummy.html.)
76. Montana Earth Science Picture of the Week.
(http://www.montanadinosaurdigs.com/brachleo.htm.)
77. Video «Meet Leonardo, The Mummy Dinosaur». Mr. Video Productions.
13.11.2005.
(http://mrvideo.com/ZenCart/index.php?main_page=product_info&products_id=1.)
78. Judith River Dinosaur Institute. 2006.
(http://formontana.net/malta.html.)
79. Fossilized dinosaur mummy gets X-rayed. A Science Museum of
Minnesota Community.
(http://ltc.smm.org/buzz/blog/fossilized_dinosaur_mummy_gets_x_rayed.)
80. LaFrance D. Amazing Dinosaur Find. Posted 3.14.2003.
(http://bhcsa.org/Essay-AmazingDinosaurFind.asp.)
81. Perkins S. Dear Mummy: Rare fossil reveals common dinosaur's soft
tissue // Science News. 2002. Vol. 162. №. 16. P. 243.
(http://www.sciencenews.org/articles/20021019/fob2.asp.)
82. Chin K., Eberth D.A., Schweitzer M.H. et al. Remarkable preservation
of undigested muscle tissue within a Late Cretaceous tyrannosaurid coprolite
from Alberta, Canada // Palaios. 2003. V. 18. № 3. P. 286–294.
83. Sutton M.D., Briggs D.E., Siveter D.J., Siveter D.J. Fossilized soft
tissues in a Silurian platyceratid gastropod // Proc. Biol. Sci. 2006. V. 273.
№ 1590. P. 1039-1044.
84. Donoghue P.C., Bengtson S., Dong X.P. et al. Synchrotron X-ray
tomographic microscopy of fossil embryos // Nature. 2006. V. 442. № 7103. P.
680–683.
85. Ashworth A.C., Thompson F.Ch. Palaeontology: A fly in the
biogeographic ointment // Nature. 2003. V. 423. № 6936. P. 135–136.
86. Perkins S. Winging South: Finally, a fly fossil from Antarctica // Science News Online. 2003. V. 163. № 19. P. 292. (http://www.sciencenews.org/articles/20030510/fob3.asp.)
87. Веденеева Н. Сенсация высокого полета: ученые откопали под Волгоградом останки древнейшей птицы // МК. 27.12.2006. С. 1, 3. (Есть сетевая версия.)
88. Курочкин Е.Н. Список публикаций (1971–2006) // Палеонтологический институт РАН. (http://www.paleo.ru/institute/publications.html?author=%CA%D3%D0%CE%D7%CA%C8%CD%2B%C5.%CD.)
89. Jones T.D., Ruben J.A., Martin L.D., Kurochkin E.N., Feduccia A. et
al. Nonavian feathers in a late Triassic archosaur // Science. 2000. V. 288. №
5474. P. 2202–2205.
90. Виланд К. Камни и
кости. Симферополь: Паломник.
2000. — 48 с.
91.
Armstrong W.G., Halstead L. B., Reed F.B., Wood L. Fossil Proteins in
Vertebrate Calcified Tissues // Philos. Trans. R. Soc. Lond. B. Biol. Sci.
1983. V. 301. № 1106. P. 301–343.
92.
Bada J.L., Wang X.S., Hamilton H. Preservation of key biomolecules in the
fossil record: current knowledge and future challenges // Philos. Trans. R.
Soc. Lond. B. Biol. Sci. 1999. V. 354. № 1379. P. 77–86.
93.
Gibbons A. Plucking the Feathered Dinosaur // Science. V. 278.№ 5341. P.
1229–1230. Dinosaur discoveries in Patagonia, Madagascar, and China rekindled
debate about the hypothesis that birds evolved from dinosaurs // Science News.
1997. V. 152. № 20. P. 310.
94.
Dinosaur Fossil Bone Leads to Gender, Age Determinations // Site NC State University
News Services. Tracey Peake, News Services, 2/06/2005.
(http://www.ncsu.edu/news/press_releases/05_06/133.htm.)
95.
Feduccia A, Nowicki J.The hand of birds revealed by early ostrich embryos //
Naturwissenschaften. 2002. V. 89. № 9. P. 391–393.
96.
Williamson D. Scientist says ostrich study confirms bird 'hands' unlike those
of dinosaurs // University Of North Carolina. UNC News Services. 2002. New 425.
(http://www.unc.edu/news/newsserv/research/feduccia081402.htm;
http://www.sciencedaily.com/releases/2002/08/020815072053.htm.)
97.
Lee A.J., Hodges S., Eastell R. Measurement of osteocalcin // Am. Clin.
Biochem. 2000. V. 37. P. 4332–4446. (Есть сетевая версия.)
98. Nielsen-Marsh C.M., Ostrom P.H., Gandhi H. et al. Sequence preservation of osteocalcin protein and mitochondrial DNA in bison bones older than 55 ka // Geology. 2002. V. 30. № 12. P. 1099–1102. (Есть сетевая версия.)
99. Уайт А., Хендлер Ф., Смит Э. и др. Основы биохимии: в 3-х томах. Т. 3. Пер. с англ. под ред. Ю.А. Овчинникова. М.: Мир. 1981. — 726 с.
100. Ленинджер А. Основы биохимии.
Пер. с англ. под ред. акад. А.А. Баева и д-ра хим. наук
Я.М. Варшавского. М.: Мир,
1976. — 962 с.
101. Stu B. Prions Research ascelerates // Chem. and Eng. News. 1998. V. 76. № 6. P. 22–29.
102. Чард Т. Радиоиммунологические методы. Пер. с англ. под ред. Я.М. Варшавского. М.: Мир. 1981. — 248 с.
103. Ткачева Г.А., Балаболкин М.И., Ларичева И.П. Радиоиммунохимические методы исследования. Справочник. М.: Медицина, 1983. — 192 с.
104. Иммунология. Под ред. У. Пола. В трех томах. Т. 3. Пер. с англ. М.: Мир, 1989. — 360 с.
105. Галактионов В.Г. Иммунология. М.: Нива России, 2000. — 488 с.
106. Schweitzer M.H., Watt J.A., Avci R. et al. Keratin immunoreactivity
in the Late Cretaceous bird Rahonavis
ostromi // J. Vert. Paleontology. 1999. V. 19. № 4. P. 712–722.
107. Borja C., Garcia-Pacheco M., Olivares E.G. et al. Immunospecificity
of albumin detected in 1.6 million-year-old fossils from Venta Micena in Orce,
Granada, Spain // Am. J. Phys. Anthropol. 1997. V. 103. № 4. P. 433–441.
108. Gibert J., Campillo D., Arques J.M. et al. Hominid status of the
Orce cranial fragment reasserted // J. Human Evolution. 1998. V. 34. P.
203–217. (Есть сетевая версия.)
109. Lowenstein J.M. Fossil proteins and evolutionary time // Pontif.
Acad. Sci. Scr. 1983. V. 50. P. 151–162.
110. Rowley M.J., Rich P.V., Rich T.H., Mackay I.R. Immunoreactive
collagen in avian and mammalian fossils // Naturwissenschaften. 1986. V. 73. P.
620–623.
111. Wick G., Kalischnig G., Maurer H. et al. Really
old-palaeoimmunology: immunohistochemical analysis of extracellular matrix
proteins in historic and pre-historic material // Exp. Gerontol. 2001. V. 36. №
9. P. 1565–1579.
112.
Schweitzer M.H., Cano R.J., Starkey J.R., Horner J.R. Multiple lines of
evidence for the preservation of collagen and other biomolecules in
unmineralized bone from Tyrannosaurus rex
// J. Vert. Paleontol. 1994. V. 14. Sup. 3. P. 45A.
113.
Schweitzer M.H., Wittmeyer J.L., Horner J.R., Toporski J.K. Soft-Tissue Vessels
and Cellular Preservation in Tyrannosaurus
rex. 2005. Supporting Online Material
(www.sciencemag.org/cgi/content/full/307/5717/1952/DC1. В 2005 г. была сетевая
версия; в 2006 г. стала недоступна.)
114.
Avci R., Schweitzer M.H., Boyd R.D. et al. Preservation of bone collagen from
the late Cretaceous period studied by immunological techniques and atomic force
microscopy // Langmuir. 2005. V. 21. № 8. P. 3584–3590.
115.
Rothschild B. Contributions of paleorheumatology to understanding contemporary
disease // Reumatismo. 2002. V. 54. № 3. P. 272–284.
116.
Gurley L.R., Valdez J.G., Spall W.D. et al. Proteins in the fossil bone of the
dinosaur, Seismosaurus // J. Protein Chem.
1991. V. 10. № 1. P. 75–90.
117.
Wyckoff R.W.G. The Biochemistry of Animal Fossils. Bristol: Scientechnica.
1972.
118.
Muyzer G., Sandberg P., Knapen M.H.J. et al. Preservation of the bone protein
osteocalcin in dinosaurs // Geology. 1992. V. 20. P. 871–874.
119. Monastersky R. Protein identified in dinosaur fossils — osteocalcin
could not be isolated in research, but may be oldest protein // Science News,
Oct 3, 1992.
http://www.findarticles.com/p/articles/mi_m1200/is_n14_v142/ai_12735009.
120. Dinosaur Blood and DNA //
http://www.ridgenet.net/~do_while/sage/v3i8f.htm.
121.
Embery G., Milner A.C., Waddington R.J. et al. Identification of proteinaceous
material in the bone of the dinosaur Iguanodon
// Connect. Tissue Res. 2003. V. 44. Suppl. 1. P. 41–46.
122. Torres J.M., Borja C., Olivares E.G.
Immunoglobulin G in 1.6 Million-year-old Fossil Bones from Venta Micena
(Granada, Spain) // J. Archaeol. Sci. 2002. V. 29. № 2. P. 167–175.
123. Embery G., Milner A., Waddington R.J. et al. The isolation and detection
of non-collagenous proteins from the compact bone of the dinosaur Iguanodon // Connect. Tissue Res.
2000.V. 41. № 3. P. 249–259.
124. Lowenstein J.M., Scheuenstuhl G., Eglinton G. et al. Immunological
methods in molecular palaentology // Phil. Trans. Roy. Soc. Lond. B. 1991. V.
333. № 1268. P. 375–380.
125.
Curry G.B. Molecular palaeontology. In Briggs, D.E.G. and Crowther, P.R. (eds),
Palaeobiology: A Synthesis. Blackwell Scientific Publications, Oxford. 1990. P.
95–100.
126.
Tuross N. The biochemistry of ancient DNA in bone // Experientia. 1994. V. 50.
№ 6. P. 530–535.
127. Tuross N., Stathoplos L. Ancient proteins in fossil bones //
Methods in Enzymology. 1993. V. 224. P. 121–129.
128. Canfield D.E., Raiswell R. Carbonate precipitation and dissolution.
In Allison P.A. and Briggs D.E.G. (eds), Taphonomy: Releasing the Data Locked
in the Fossil Record. Plenum Press, NY. 1991. — P. 411–453.
129. Ambler R.P., Daniel M. Proteins and molecular palaeontology //
Philos. Trans. R. Soc. Lond. B. Biol. Sci. B. 1991. V. 333. № 1268. P. 381–389.
130. Logan G., Collins M.J., Eglinton J. Preservation of organic
biomolecules. In Allison P.A. and Briggs D.E.G. (eds), Taphonomy: Releasing the
Data Locked in the Fossil Record. Plenum Press, NY. 1991. — P. 1–24.
131. Weiner S., Traub W., Elster H., DeNiro M.J. The molecular structure
of bone and its relation to diagenesis // Appl. Geochem. 1989. V. 4. № 3. P.
231–232.
132. Sykes G.A., Collins M.J., Walton D.I. The significance of a
geochemically isolated intracrystalline organic fraction within biominerals //
Organic Geochemistry. 1995. V. 23. № 11–12. P. 1059–1065.
133. Smith A.J., Matthews J.B., Wilson C., Sewell H.F. Plasma proteins
in human cortical bone: in vitro binding studies // Calcif. Tissue Int. 1985.
V. 37. № 2. P. 208–210.
134. Berman A., Addadi L., Weiner S. Interactions of sea-urchin
macromolecules with growing calcite crystals — a study of intracrystalline
proteins // Nature. 1988. V. 331. № 6156. P. 546–548.
135. Norris S. Many Dino Fossils Could Have Soft Tissue Inside //
National Geographic News. February 22, 2006.
(http://news.nationalgeographic.com/news/2006/02/0221_060221_dino_tissue.html.)
136. Paleontologist presents theories of fossil preservation at AAAS //
Bulletin Online. News for the North Carolina State University Community.
02.24.2006. (http://www.ncsu.edu/BulletinOnline/02_06/AAASfollow.htm.)
137. Rotilio G. Biochemical mechanisms of oxy-radical production and the role of the antioxidant enzymes. In: Oxygen Free Radicals in Shock. Int. Workshop, Florence, 1985. Ed. by N. Ursini, Karger, Basel, 1986. P. 1–8.
138. Новиков К.Н. Перекисное
окисление в гепатоцитах. В кн.: Гепатоцит: Функционально-метаболические
свойства. М.: Наука. 1985. С.
146–169.
139. Wade C.R., Van Rij A.M. Plasma thiobarbituric acid reactivity:
reaction condition and the role of iron, antioxidants and lipid peroxy radicals
on the quantitation of plasma lipid peroxides // Life Sci. 1988. V. 43. № 13.
P. 1085–1093.
140. Kirkpatrick D.T., Guth D.J., Mavis R.D. Detection of in vivo lipid
peroxidation using the thiobarbituric acid assay for lipid hydroperoxides // J.
Biochem. Toxicol. 1986. V. 1. P. 93–104.
141. Gutteridge J.M.C. Free radical damaged to lipids, amino acid,
carbohydrates and nucleic acids determined by thiobarbituric acid reactivity //
Int. J. Biochem. 1982. V. 14. № 7. P. 649–653.
142. Lowenstein J.M., Sarich V.M., Richardson B.J. Albumin systematics
of the extinct mammoth and Tasmanian wolf // Nature. 1981. V. 291. № 5814. P.
409–411.
143. Cattaneo C., Gelsthorpe K., Phillips P., Sokol R.J. Blood in ancient
human bone // Nature. 1990. V. 347. № 6291. P. 339.
144. Cattaneo C., Gelsthorpe K., Phillips P., Sokol R.J. Reliable
identification of human albumin in ancient bone using ELISA and monoclonal
antibodies // Am. J. Phys. Anthropol. 1992. V. 87. № 3. P. 365–372.
145. Cattaneo C., Gelsthorpe K., Phillips P. Sokol R.J. Blood residues
on stone tools: indoor outdoor experiments // World Archaeol. 1993. V. 25. P.
20–45.
146. Brandt E., Wiechmann I, Grupe G. How reliable are immunological
tools for the detection of ancient proteins in fossil bones? // Int. J.
Osteoarchaeology. 2002. V. 12. № 5. P. 307–316.
147. Downs E.F., Lowenstein J.M. Identification of archaeological blood
proteins: a cautionary note // J. Archaeol. Sci. 1995. V. 22. № 1. P. 11–16.
148. Cattaneo C., Gelsthorpe K., Phillips P., Sokol R.J. Detection of
human proteins in buried blood using ELISA and monoclonal antibodies: towards
the reliable species indentification of blood stains on buried material //
Forensic Sci. Int. 1992. V. 57. № 2. P. 139–146.
149. Schmidt-Schultz T.H., Schultz M. Bone protects proteins over
thousands of years: extraction, analysis, and interpretation of extracellular
matrix proteins in archeological skeletal remains // Am. J. Phys. Anthropol.
2004. V. 123. № 1. P. 30–39.
150. Lendaro E., Ippoliti R., Bellelli A. et al. Brief communication. on
the problem of immunological detection of antigens in skeletal remains // Am.
J. Phys. Anthropol. 1991. V. 86. № 3. P. 429–432.
151. Smith P.R., Wilson M.T. Detection of haemoglobin in human skeletal
remains by ELISA // J. Arch. Sci. 1990. V. 17. P. 255–268.
152. Nielsen-Marsh C.M., Richards M.P., Hauschkad P.V. et al.
Osteocalcin protein sequences of Neanderthals and modern primates // Proc.
Natl. Acad. Sci. U.S.A. 2005. V. 102. № 12. P. 4409–4413. (Есть сетевая версия.)
153. Monsalve M.V., Stone A.C., Lewis C.M. et al. Brief communication:
molecular analysis of the Kwaday Dan Ts'finchi ancient remains found in a
glacier in Canada // Am. J. Phys. Anthropol. 2002. V. 119. № 3. P. 288–291.
154. Perrin C., Noly V., Mourer R., Schmitt D. Preservation of cutaneous
structures of egyptian mummies. An ultrastructural study // Ann. Dermatol.
Venereol. 1994. V. 121. № 6–7. P. 470–475.
155. Nerlich A.G., Parsche F., Kirsch T. et al. Immunohistochemical detection
of interstitial collagens in bone and cartilage tissue remnants in an infant
Peruvian mummy // Am. J. Phys. Anthropol. 1993. V. 91. № 3. P. 279–285.
156. Grupe G., Turban-Just S. Amino acid composition of degraded matrix
collagen from archaeological human bone // Anthropol. Anz. 1998. V. 56. № 3. P.
213–226.
158. Bocherens H., Grupe G., Mariotti A., Turban-Just S. Molecular
preservation and isotopy of Mesolithic human finds from the Ofnet cave
(Bavaria, Germany) // Anthropol. Anz. 1997. V. 55. № 2. P. 121–129.
159. Schaedler J.M., Krook L., Wootton J.A. et al. Studies of collagen
in bone and dentin matrix of a Columbian mammoth (late Pleistocene) of central
Utah // Matrix. 1992. V. 12. № 4. P. 297–307.
160. Franc S., Marzin E., Boutillon M.M. et al. Immunohistochemical and
biochemical analyses of 20,000–25,000-year-old fossil cartilage // Eur. J.
Biochem. 1995. V. 234. № 1. P. 125–131.
161. Palmqvist P. A critical reevaluation of the evidence for the
presence of hominids in lower pleistocene times at Venta-Micena, southern Spain
// J. Hum. Evolution 1997. V. 33. № 1. P. 83–89.
162. Wieland C. Sensational dinosaur blood report! // Creation. 1997. V.
19. № 4. P. 42–43. (http://www.answersingenesis.org/docs/4232cen_s1997.asp.)
163. Yeoman B. Schweitzer’s Dangerous Discovery // Discover. 2006. V.
27. № 4. P. 37-41.
(http://www.discover.com/issues/apr-06/features/dinosaur-dna/?page=1.)
164. Wieland C. Dinosaur bones: Just how old are they really? An
evolutionary dinosaur expert reveals some fascinating facts! // Creation. 1999.
V. 21. № 1. P. 54–55. (http://www.answersingenesis.org/docs/3947.asp.)
165. Wieland C. Evolutionist questions AiG report — Have red blood
cells really been found in T. rex fossils?
// 2002. (http://www.answersingenesis.org/docs2002/0325rbcs.asp.)
166. Hurd G.S. Dino-blood and the Young Earth // Talk Origin Archive.
2004. (http://www.talkorigins.org/faqs/dinosaur/blood.html;
http://home.austarnet.com.au/stear/YEC_and_dino_blood.htm.)
167. Catchpoole D., Sarfati J. Schweitzer’s Dangerous Discovery //
Creation Ministries International — formerly Answers in Genesis. 19 July
2006. (http://www.creationontheweb.com/content/view/4504.)
168. Lovett R.A. Fossils Yield 10-Million-Year-Old Bone Marrow — A
First // National Geographic News. July 25, 2006.
(http://news.nationalgeographic.com/news/2006/07/060725-fossil-bone.html.)
169. Hurd G.S. Ancient Molecules and Modern Myths // Talk Origin
Archive. 2004. (http://www.talkorigins.org/faqs/dinosaur/osteocalcin.html.)
170. Dinosaur. From Wikipedia, the free encyclopedia. 2005
(http://en.wikipedia.org/wiki/Dinosaur.)
171. Stokstad E. Tyrannosaurus rex Soft Tissue Raises Tantalizing Prospects //
Science. 2005. V. 307. № 5717. P. 1852. (Есть сетевая версия.)
172. Schweitzer M.H., Wittmeyer J.L., Horner
J.R. Soft tissue and cellular preservation in vertebrate skeletal elements from
the Cretaceous to the present // Proc. Biol. Sci. 2007. V. 274. № 1607. P.
183–197.
173. McNamara M.E., Orr P.J., Kearns S.L. et
al. High-fidelity organic preservation of bone marrow in ca. 10 Ma amphibians
// Geology. 2006. V. 34. №. 8. P. 641–644.
174.
Fossil frogs yield 'soft tissues' // BBC. July 26, 2006.
(http://www.crystalinks.com/fossilfrogs.html.)
175.
Bone marrow still juicy after 10 million years // New Scientist. 2006. № 2563.
P. 17.
(http://www.newscientist.com/channel/life/mg19125634.900-bone-marrow-still-juicy-after-10-million-years.html.)
176. Козинец Г.И., Неменова Н.М., Терентьева
Э.И. и др. Костный мозг. Большая медицинская энциклопедия. 3-е издание. Под
ред. акад. Б.В. Петровского. Т. 11. М.: Сов. энциклопедия, 1979. С.
435–441.
177.
Frog fossil spawns ancient bone-marrow find // Independent Online. August 02
2006.
(http://www.iol.co.za/index.php?set_id=1&click_id=31&art_id=qw1154535304642B264/)
178.
Ker Th.K. Bone Marrow Found Intact in Ancient Fossils // Live Science. July 26,
2006. (http://www.foxnews.com/story/0,2933,205515,00.html.)
179.
Mayell H. T. Rex Soft Tissue Found
Preserved // National Geographic News. March 24, 2005.
(http://news.nationalgeographic.com/news/2005/03/0324_050324_trexsofttissue.html.)
180.
Maat G.J.R., Baig M.S. Microscopy electron scanning of fossilized sickle-cells
// Int. J. Anthropology. 1990. V. 5. № 3. P. 271–275.
181. Bliss F. (MS Biostratigraphy). Letter about fossilized bone marrow. 7.27.06. (http://dml.cmnh.org/2006Jul/msg00390.html.)
182. Еремин Н.И. Неметаллические полезные ископаемые. М.: Изд-во МГУ, 2004. (http://geo.web.ru/db/msg.html?mid=1172887&uri=glava_2.htm.)
183. Spanish minerals. Mineral Specimen No. 00006: Sulfur from Cantabria, Spain. (http://www.spanishminerals.com/00006.htm.)
184. Полинг А. Общая химия. Пер с англ. В.М.
Сахарова. Под ред. проф. М.Х.
Карапетьянца. М.:
Мир, 1974. — 848 c.
185.
Ancient DNA answers questions from the past // The University of Queensland. UQ
News Online. 26 May 2004. (http://www.uq.edu.au/news/index.html?article=5569;
www.brisinst.org.au/people/loy_tom.html.)
186.
UQ researcher unlocks two-million-year-old lifestyle // The University of
Queensland. UQ News Online. 30 September 1998.
(http://www.uq.edu.au/news/index.html?article=1005.)
187.
In Memoriam. Thomas Harold Loy, 1942–2005 // Personal homage brother Dr. Gareth
Loy. (http://www.garethloy.com/TomLoy/In_Memoriam_Tom_Loy.html.)
188.
Loy T.H. Prehistoric blood residues: detection on tool surfaces and identification
of species of origin // Science. 1983. V. 220. № 17. P. 1269–1271.
189.
Loy T.H. Recent advances in blood residue analysis // In: Ambrose W.R., and
Mummer J.M.J. (eds), Archaeometry: further Australasian Studies. 1987.
Australian National University, Canberra. P. 57–65.
190.
Loy T.H., Wood A.R. Blood residue analysis at Cayonu Tepesi, Turkey // J. Field
Archaeology. 1989. V. 16. P. 451–460.
191.
Loy T.H., Hardy B.L. Blood residue analysis of 90,000 year-old stone tools from
Tabun Cave, Israel // Antiquity. 1992. V. 66. № 250. P. 24–35.
192.
Loy T.H. The artifact as site: an example of the biomolecular analysis of
organic residues on prehistoric tools // World Archaeol. 1993. V. 25. № 1. P.
44–63.
193.
Loy T.H., Dixon E.J. Blood Residues on Fluted Points from Eastern Beringia //
Amer. Antiquity. 1998. V. 63. № 1. P. 21–46.
194.
Kaiser J. Blood from Stones. Tests for prehistoric blood cast doubt on earlier
results // Science News. 1995. V. 147. № 24. P. 376. (Есть сетевая версия.)
195.
Salleh A. Bloody stone tools tell hominids' tales // ABC Science Online. News
in Science. 19 July 2004. (http://www.abc.net.au/science/news/stories/s1156792.htm.)
196.
7th International Conference on Ancient DNA and Associated Biomolecules, 2004.
Brisbane, Australia. (http://www.uq.edu.au/news/index.html?article=5569.)
197.
Using new tools on old weapons (Dr. Thomas Loy) // Catalyst Queensland. 2004. №
9. P. 6–7. (Есть сетевая версия.)
198.
Lowenstein J.M. Molecular Evolution and the Fossil Record: Short Courses in
Paleontology, ed. Broadhead, T.W. (Paleontological Society), 1988. P. 12–19.
199. Otzi. The Iceman // Ellie Crystal’s Metaphysical and Science Website. (http://www.crystalinks.com/oetzi.html.)
200. Месть воина Ледникового периода // Мир
путешествия и приключений. 07.11.2005. (http://www.outdoors.ru/news/2005/11/07_4.php.)
201.
Bada J., Kminek G. Ionizing radiation and the long-term survival of
biomolecules and viable bacteria // Philadelphia Annual Meeting, 22–25 October
2006. Session № 125. Evidence for Long-Term Survival of Microorganisms and Preservation
of DNA. The Geological Society of America. 2006. P. 125–128.
(http://gsa.confex.com/gsa/2006AM/finalprogram/abstract_114780.htm.)
202.
Astrobiology. Published articles // Berkeley.edu. Solar system exploration.
2004. (http://astrobiology.berkeley.edu/science.htm.)
203. Kminek G., Bada J.L., Pogliano K., Ward J.F. Radiation-Dependent Limit for the Viability of Bacterial Spores in Halite Fluid Inclusions and on Mars // Radiat. Res. 2003. V. 159. P. 722–729.
204. Dinosaur Bones & Fossils // Scholastic Inc. 2007–1996. (http://content.scholastic.com/browse/article.jsp?id=4738.)
205. Алифанов В.Р., Курочкин Е.Н.,
Забелин В.И., Кудрявцев В.И. Первые находки динозавров в Туве // Природа. 2002.
№ 2. (http://www.courier.com.ru/priroda/pr0202new15.htm.)
206. Kuban G. Dangers of Rock Dust
(http://www.gemcutters.org/LDA/RockDustDangers.htm.)
207. Radiant bones. In: Salmitie M. A journey to dinosaur country // Universitas Helsingiensis. 2001. (http://www.helsinki.fi/lehdet/uh/201i.htm.)
208. Лескова Н. Динозавры вымерли от
лейкемии (В костях вымерших гигантов порой находят настоящие «урановые
рудники») // Независимая газета. Наука. 26.03.2003.
(http://www.ng.ru/science/2003-03-26/11_dinosaurus.html.)
209. Discovering Cedar Mountain Dinosaurs.
(https://scientists.dmns.org/sites/kencarpenter/Cedar%20Mountain%20storage/History.aspx?PageView=Shared.)
210. Dino finder merits patent // Continuum. The Magazine of the
University of Utah. 1998. V. 7. № 3.
(http://www.alumni.utah.edu/continuum/winter97/gazette.html.)
211. Blake E. Shades of Redwood. Hot Fossils // The Magazine of the
California Academy of Sciences. 1999. V. 52. № 1.
(http://www.calacademy.org/calwild/1999winter/stories.)
212. E. Ray Garton resume and publication. Prehistoric West Virginia.
(http://www.prehistoricplanet.com/wv/garton.htm.)
213. Wilson M.L. Radioactivity specimens in museum collections // The
Society of Mineral Museum Professionals. 2006.
(http://www.agiweb.org/smmp/rept-rad-cmnh.htm.)
214. Chure D. Fossil Vertebrates As Radon Sources: Health Update //
Conserve O Grame. National Park Service (USA). 1993. № 2/5.
(http://www.cr.nps.gov/museum/publications/conserveogram/02-05.pdf.)
215. Hall K. Storage Concerns For Geological Collections // Conserve O
Grame. National Park Service (USA). 1998. № 11/2.
(http://www.cr.nps.gov/museum/publications/conserveogram/11-02.pdf.)
216. Jiggens T.E., Cardarelli J.J., Ahrenholz S.H. et al. Health Hazard
Evaluation (HHE) report and recommendations (HETA 96-0264-2713) // Hagerman
Fossil Beds National Monument, National Park Service, U.S. Department of the
Interior Hagerman, Idaho. September 1998. — 52 p.
(http://www.cdc.gov/niosh/hhe/reports/pdfs/1996-0264-2713.pdf.)
217. Bowie S.H.U. An Unusually Radioactive Fossil Fish from Thurso,
Scotland // Nature. 1966. V. 211. P. 728–729.
218. Madson M.E. Radioactive Turtle Shells // Rocks and Minerals 1983.
V. 58. P. 245–246.
219. Khandelwal G.S., Singh J.J. Radioactivity of whale vertebra fossils
in the Chesapeake Bay area // Health Phys. 1978. V. 35. № 2. P. 411–413.
220. Singh N.P., Singh B., Singh K., Virk H.S. Autoradiographic study of
uranium and thorium in fossil bones // Int. J. Radiat. Appl. Instrum. E. Nucl.
Geophys. 1992. V. 6. № 2. P. 287–291.
221. Cloos E. Articles on Literature on Uranium and Thorium and
Radioactive Occurrences in the United States, A New Jurassic Dinosaur from
Kayenta Formation of Arizona, and Index // Bulletin Geological Society of
America. 1954. V. 65. № 6. P. 467–603.
222. Gillette D.D. Seismosaurus: The Earth Shaker. Columbia University
Press, New York, 1994. — 205 p. (http://findarticles.com/p/articles/mi_m0GDX/is_1997_May/ai_53289154.)
223. Jaworowsky Z., Pensko J. Unusually Radioactive Fossil Bones from
Mongolia // Nature. 1967. V. 214. P. 161–163.
224. Gun-Aazhav T., Gerbish Sh., Otgonsuren O., Sereeter Zh. and D. Chultem. Investigation of the radioactivity of dinosaur bones with a high-resolution gamma spectrometer // Atomic Energy. Springer New York. 1973. V. 35. № 2. P. 755–757.
225. Vasidov A., Osinskaya N.S., Khatamov Sh. Kh. et al. The neutron
activation analyses of the ancient bone remains // 5th Conference on Nuclear
and Particle Physics, Cairo, Egypt, 19–23 Nov. 2005. AR1-3.
(www.geocities.com/Athens/Library/7348/NUPPAC_05abstb.doc.)
226. Vasidov A., Mamadalieva U., Shaakhmedov А. The study of activity of
the radon gases from bone remains with detectors CR-39 // 5th Conference on
Nuclear and Particle Physics, Cairo, Egypt, 19–23 Nov. 2005. RMD2-1.
(www.geocities.com/Athens/Library/7348/NUPPAC_05abstb.doc.)
227. Butts J.J., Katz R. Theory of RBE for heavy ion bombardment of dry
enzymes and viruses // Radiat. Res. 1967. V. 30. № 4. P. 855–871.
228. Malinen E., Hult E.A., Hole E.O., Sagstuen E. Alanine radicals, part
4: relative amounts of radical species in alanine dosimeters after exposure to
6–19 MeV electrons and 10 kV–15 MV photons // Radiat. Res. 2003. V. 159. № 2.
P. 149–153.
229. Puchala M., Szweda-Lewandowska Z., Kiefer J. The influence of
radiation quality on radiation-induced hemolysis and hemoglobin oxidation of
human erythrocytes // J. Radiat. Res. (Tokyo). 2004. V. 45. № 2. P. 275–279.
230. Dartnell L.R., Desorgher L., Ward J.M., Coates A.J. Martian
sub-surface ionising radiation: biosignatures and geology // Biogeosciences
Discuss. 2007. V. 4. P. 455–492. (www.biogeosciences-discuss.net/4/455/2007/)
231. Kempner E.S., Haigler H.T. The Influence of Low Temperature on the Radiation Sensitivity of Enzymes // J. Biol. Chem. 1982. V. 255. № 22. P. 13297–13299. (Есть сетевая версия.)
232. Beliveau R., Demeule M., Jette M., Potier M. Molecular sizes of amino acid transporters in the luminal membrane from the kidney cortex, estimated by the radiation-inactivation method // Biochem. J. 1990. V. 268. № 1. P. 195–200. (Есть сетевая версия.)
233. Le Maire M., Thauvette L., De
Foresta B. et al. Effects of ionizing radiations on proteins. Evidence of
non-random fragmentations and a caution in the use of the method for
determination of molecular mass // Biochem. J. 1990. V.
267. P. 431–439. (Есть сетевая версия.)
233а. Schweitzer M.H., Suo Z., Avci R. et al.
Analyses of soft tissue from Tyrannosaurus
rex suggest the presence of protein // Science. 2007. V. 316. № 5822. P.
277–280.
233б.
Asara J.M., Schweitzer M.H., Freimark L.M. et al. 2007, Protein sequences from
mastodon and Tyrannosaurus rex
revealed by mass spectrometry / Science. 2007. V. 316. № 5822. P. 280–285.
233в.
Perkins S. Ancient Extract: T. rex
fossil yields recognizable protein // Science Online. (http://www.sciencenews.org/articles/20070414/fob3ref.asp.)
233г.
Rincon P. Protein links T. rex to
chickens // Science reporter, BBC News
(http://news.bbc.co.uk/2/hi/science/nature/6548719.stm.)
233д.
Карпов М. Белок доказывает родство королевского тираннозавра с современной курицей
// TerraLab. 16.04.2007
(http://www.terralab.ru/news/315163/.)
233е.
DeWitt D.A. Tyrannosaurus rex: a big
chicken? // Answers in Genesis. April 13, 2007.
(http://www.answersingenesis.org/articles/2007/04/13/t-rex-big-chicken.)
233ж.
T. rex related to chickens //
(http://www.livescience.com/animalworld/070412_dino_tissues.html.)
233з. Nielsen-Marsh C. Biomolecules in fossil remains. Multidisciplinary approach to endurance // The Biochemist (Journal of The Biochemical Society). June 2002. P. 12–14. (Есть сетевая версия.)
234. Цыпин Леонид, священник. Так чем же являются Дни Творения? Центральная проблема экзегетики Шестоднева. Киев: Пролог. 2005.
235. Цыпин Леонид, священник. Вера и наука. Правы ли современные «креационисты»? // Сайт «Киевская Русь». (Kiev-orthodox.org.)
236. О том, что препятствует деятельности Церкви и государства в сфере образования (интервью с диаконом Андреем Кураевым) // Царкоунае слова (Минск). 2006. № 4; аналогичный материал: сайт Крутицкого подворья (krutitsy.ru) и сайт «Интерфакс-религия» (www.interfax-religion.ru/ortodoxy).
237. Муравник Г.Л. Ибо прах ты и в прах возвратишься. // Христианство и наука. Сб. докл. конф. Х Международных образовательных чтений. Москва. 2003.
238. Муравник Г.Л. Ни шагу вперед! // Полемика об эволюции. (http://evolution.powernet.ru/polemics/vertyanov.htm.)
239. «Той повеле и создашася». Современные ученые о сотворении мира (Сборник). Клин: Фонд «Христианская жизнь». 1999. — 192 с.
240. Борисов Н.М. (Институт ядерной безопасности и защиты, Франция). Христианский взгляд на мироздание или распространение слухов? Отзыв на книги по «креационной науке» издательства «Паломникъ». (http://evolution.powernet.ru/polemics/borisov.html.)
241. Серафим (Роуз), иеромонах. Православный взгляд на эволюцию // Приношение православного американца. М., 1998. (Есть сетевые версии.)
242. Серафим (Роуз), иеромонах. Православное понимание книги Бытия. М., 1998. (Есть сетевые версии.)
243. Буфеев Константин, священник (составитель). Свидетельства Святых Отцов против эволюции [62 свидетельства]. В кн.: «Православное осмысление мiра». Материалы XIII международных рождественских образовательных чтений. «Шестодневъ». М., 2005. С. 10–35. (Есть сетевая версия.)
244. Буфеев Константин, священник (составитель). Свидетельства святых о церковной вере в шестидневное творение Богом мiра [157 свидетельств]. В кн.: «Православное осмысление творения мiра». Выпуск 2. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», Москва, 2006. С. 6–44. (Есть сетевая версия.)
245. Буфеев Константин, священник (составитель). Молитвенные тексты из Октоиха, свидетельствующие о церковном отношении к Адаму и Еве как к реальным историческим личностям, созданным Богом в шестой день творения. В кн.: «Православное осмысление творения мiра». Выпуск 3. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», М.: Изд-во «Шестодневъ», 2007. С. 11–26. (Есть сетевая версия.)
246. Буфеев Константин, священник. Православное вероучение и теория эволюции. С.-Пб.: Общ-во святителя Василия Великого. 2003. — 496 с. (Есть сетевая версия.)
247. Буфеев Константин, священник. О церковной оценке тейярдизма и о прославлении иеромонаха Серафима (Роуза) в лике святых Русской Православной Церкви. О ереси эволюционизма. В кн.: «Православное осмысление мiра». Материалы XIII международных рождественских образовательных чтений. «Шестодневъ». М., 2005. С. 36–61. (Есть сетевая версия.)
248. Буфеев Константин, священник. «Православный эволюционизм» как одно из новых «мнений Церкви». В кн.: «Православное осмысление творения мiра». Выпуск 3. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», М.: Изд-во «Шестодневъ», 2007. С. 55–80. (Есть сетевая версия.)
249. Буфеев Константин, священник. Отец Александр Мень: сказочник или сочинитель апокрифов? В кн.: «Православное осмысление творения мiра». Выпуск 3. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», М.: Изд-во «Шестодневъ», 2007. С. 370–396. (Есть сетевая версия.)
250. Сысоев Даниил, диакон. Летопись начала. Изд-ние Сретенского монастыря, 1999. — 256 с. (Есть сетевая версия.)
251. Сысоев Даниил, священник. Православные креационисты и эволюционисты: шесть лет спустя. В кн.: «Православное осмысление мiра». Материалы XIII международных рождественских образовательных чтений. «Шестодневъ». М., 2005. С. 118–133. (Есть сетевая версия.)
252. Священник Тимофей. Православие и современное естествознание. М.: Паломникъ. 1998. — 208 с.
253. Иеромонах Дамаскин (Христенсен). Иеромонах Серафим (Роуз) как выразитель святоотеческого отношения к вопросу об эволюции. В кн.: «Православное осмысление мiра». Материалы XIII международных рождественских образовательных чтений. «Шестодневъ». М., 2005. С. 69–82. (Есть сетевая версия.)
254. Иеромонах Дамаскин (Христенсен). Спасение в свете Творения. В кн.: «Православное осмысление творения мiра». Выпуск 2. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», Москва, 2006. С. 65–84. (Есть сетевая версия.)
255. Хоменков А.С. Психологические предпосылки эволюционного мышления. В кн.: «Православное осмысление творения мiра». Выпуск 3. Отдел религиозного образования и катехизации Русской Православной Церкви. Миссионерско-Просветительский Центр «Шестодневъ», М.: Изд-во «Шестодневъ», 2007. С. 215–223. (Есть сетевая версия.)
256. Колчуринский Н.Ю. Деятельность православного миссионера в молодежной аудитории и проблемы происхождения мiра. В кн.: «Православное осмысление мiра». Материалы XIII международных рождественских образовательных чтений. «Шестодневъ». М., 2005. С. 296–304. (Есть сетевая версия.)
(Данная статья готовится к напечатанию в альманахе «Божественное откровение и современная наука» №3)
[1] Автор приносит благодарность канд. психол. наук Н.Ю. Колчуринскому (центр «Шестодневъ»), инициировавшему начало исследований путем предоставления соответствующих ссылок. Без этой его инициативы ничего, вероятно, и не было бы.
[2] «Обыденно-научное представление» — сложившийся в сознании научного работника штамп о том, в чем он конкретно не разбирается, но думает, что разбирается, поскольку данный предмет лежит близко от его профессиональной области.
[3] «Молекулярно-клеточная палеонтология» — наименование, введенное, исходя из открывшихся реалий, автором представленного труда только в январе 2007 г. Ни в каких более ранних публикациях (в том числе и в публикациях самого автора) оно не встречается, всюду дисциплина определяется более узко, как «молекулярная палеонтология».
[4] Дисциплину, называемую «палеогенетика», которая изучает древние и ископаемые ДНК, мы к молекулярно-клеточной палеонтологии не причисляем, поэтому огромный массив соответствующих обзоров и экспериментальных статей остается пока вне рамок нашего исследования (кратко о палеогенетике см. также ниже).
[5] В [6–8] изучение ископаемых ДНК тоже относят к молекулярной палеонтологии, не вычленяя палеогенетику. По нашему мнению, это неправомерно.
[6] Доктор P.H. Abelson умер в 2004 г. в возрасте 91 года.
[7] Если подходить с современными критериями достоверности данных в молекулярной палеонтологии [8], то нет прямых доказательств того, что источником этих аминокислот является именно та девонская рыба возрастом в «350 млн. лет».
[8] Мы представляем здесь весь список работ доктора Р. Павлички по молекулярно-клеточной палеонтологии, обнаруженных в Medline (PubMed), чтобы отдать дань уважения этому почти забытому пионеру данной дисциплины.
[9] В обычном химическом выражении — более 10 тыс. углеродных единиц.
[10] Перминерализация — проникновение минералов в район сосудистых участков кости; заполнение минералами ее открытых частей.
[11] Согласно энциклопедическому словарю, псевдоморфозы (от псевдо ... и греч.
morphe — форма) — это минеральные образования, обладающие внешней
кристаллографической формой, чуждой слагающему их веществу; образуются путем
заполнения полостей, оставшихся от выщелоченных минералов (псевдоморфозы
заполнения), или химическим замещением; при этом значительная часть вещества
замещенного минерала входит в состав замещающегося (псевдоморфозы превращения)
или выносится полностью (псевдоморфозы вытеснения).
[12] Печеночники — класс мхов. Ныне насчитывают свыше 6 тыс. видов; распространены по всему земному шару, особенно в тропиках.
[13] Кератины — белки, формирующие волосы, перья,
чешую и т.п. образования. Вследствие жесткой молекулярной структуры очень
устойчивы к внешним воздействиям. β-Кератин для
современных животных обнаружен только у рептилий и птиц (чешуя, перья) [99,
100].
[14] Коллаген. Соединительная ткань организма формирует
хрящи, сухожилия, связки, остов костей и т.д. Механическая и поддерживающая
функция этой ткани обеспечивается нерастворимыми нитями, образованными
высокополимерными соединениями коллагена — самого распространенного белка
животных. Мономеры коллагена представляют собой трехнитевые белковые «тяжи»,
которые связываются друг с другом поперечными молекулярными связями (сшивками),
образуя коллаген. Такая жесткая структура обеспечивает механическую прочность
при сопутствующей эластичности [99, 100].
[15] Остеокальцин — низкомолекулярный белок,
содержащий большое количество глутаминовой кислоты; специфичен для костей.
Жесткая структура молекулы делает белок весьма стабильным, в частности, к
нагреванию: он не полностью распадается при термообработке в течение более чем
5-ти ч при 165°C [97, 98].
[16] Ранее, по крайней мере в 2005 г., и статья в «Science», и сопутствующий ей материал online были в Интернете в свободном доступе, и мы их именно тогда и извлекли. Оказалось, однако, что ныне, в начале 2007 г., публикации в свободном доступе уже отсутствуют. Почему так, мы сказать точно не можем.
[17] Непонятно, как тогда работают разобранные в подразделе 4.3. основные механизмы сохранения органических молекул: формирование комплексов с минеральной составляющей и образование стойких неорганических внутренних кристаллов. Даже по логике ясно: если нет минерализации, то и «миллионов лет» быть не должно.
[18] Автор приносит благодарность канд. техн. наук С.П. Скворцову за любезно присланную первичную ссылку по костному мозгу амфибий. Именно он первым обратил внимание на костный мозг в «10 млн. лет».
[19] Мы не удивились бы ручному рубилу и в «десять миллионов лет».
[20] Ископаемые остатки залегают в долине, промытой рекой Малой Миссури через породы мелового периода, которые простираются от Северной Дакоты в штаты Монтана и Южная Дакота (США). Этот слой горной породы известен как Адский Ручей (Hell Creek), где и находят окаменелости Мелового периода.
[21] Правильны в данном случае размерности поглощенной дозы излучения — греи (Гр) и рады, но мы приводим в скобках более известные всем «рентгены», хотя это и размерности не поглощенной, а экспозиционной дозы.
[22] Доктор Дж. Бада давно занимается в том числе астробиологией — т.е. сохранностью биомолекул, вирусов и спор бактерий в космосе и на других планетах (Марсе) [201–203]. Поэтому для него незнание точной ситуации с радиационной инактивацией сухих биомолекул особенно непростительно.
[23] Можно сказать: «находясь в плену обыденно-научных представлений» — см. выше примечание 2.
[24] Специальный момент: в отличие от живых структур, при облучении сухих биомолекул и мертвых биоструктур относительная биологическая эффективность редкоионизирующего (рентгеновское и γ-излучение, электроны) и плотноионизирующего (α-частицы, нейтроны, протоны и пр.) излучения одинакова. То есть, радиационные эффекты применительно к объектам молекулярно-клеточной палеонтологии зависят только от поглощенной дозы (поглощенной энергии), но не от качества (природы) радиации и не от степени ионизации среды излучением [227–230].
[25] Мы надеемся, что радиационный аспект в палеогенетике послужит предметом нашего следующего креационного исследования.
[26] В данном разделе мы представляем только самые
ключевые ссылки; все остальное можно найти выше.